© Shutterstock
This is called maturation of the gonads, writes Professor Nihar Ranjan Chattopadhyay, West Bengal University of Animal and Fishery Sciences.
Most of the fishes exhibit seasonal cycle in the production of gametes. The expulsion of gametes from the body into the surrounding water is called "spawning" resulting in fertilisation. Fish spawns during a specified periods of the year which coincides with the several environmental factors that are available during the said period only. The entire period during which the gonads attain full maturity and spawning takes place in the population is called the breeding season of the species.
After spawning, gonadal recrudescence may occur in the same breeding season (multiple breeding) twice or thrice depending on the availability of certain crucial environmental factors like temperature and photoperiod ie increased daylength.
Maturity stage in the female fish
On the basis of shape, size, colour of the ovary and other histomorphological features, at least six maturity stages can be recognized.
- Resting phase (immature): The ovaries are small, thin, thread like, translucent, pale or dirty white in colour with inconspicuous vascularisation. The ovaries occupy only a small part of the body cavity and ova are not visible to the naked eye. Histologically, the ovary shows ovigerous lamellae, having nests of oogonia, and immature oocytes in the stage I and II are visible under a microscope.
- Early maturing phase: Ovaries become slightly larger, thicker, opaque and are light yellowish in colour. There is an increase in the weight of the ovary and they occupy nearly ½ of the body cavity. Histologically, oocytes in stage III and IV are present in large number.
- Advanced maturing phase: There is a further increase in the weight and volume of the ovaries, which have a deep yellow colour and occupy 2/3 to ¾ of the body cavity. Vascular supply increases and the blood capillaries become conspicuous. Immature oocytes are reduced in number while stage IV and stage V oocytes are present in large number. A few stage VI oocytes may also be seen.
- Mature or prespawning phase: The ovaries are further enlarged occupying almost the entire body cavity. They are turgid, deep yellow in colour and a large number of spherical ova are visible to the naked eye through the thin ovarian wall. The blood supply increases considerably. Both translucent and opaque ova are present and the ovaries attain their maximum weight. The fish becomes gravid due to ripe ova tucked inside, and the abdomen becomes round. The ova are not discharged till the environmental conditions become favourable. Histologically, a large number of ova in stage VII, and ripe eggs are seen in the ovary.
- Spawning phase: Overies are very much enlarged, occupying the entire body cavity. They are turgid and yellow in colour with a large number of translucent eggs. Ovarian wall is very thin, almost transparent. Eggs are present in the oviduct also, and the fish spawns a number of times during this period. The ovary is now said to be in running phase. At the beginning of this phase, ova are extruded by applying a gentle pressure on the abdomen.
- Spent phase: The ovaries are flaccid, shrunk and sac-like reduced in volume and have a dull colour. The vascular supply is reduced. Some unspawned large ova and a large number of small ova are present. Histologically, the ovary shows atretic and discharged follicles, along with stage I and II oocytes.
Seasonal changes in the ovaries of teleosts have been studied by several investigators. The spawning season can be predicted by studying the occurrence of mature fish as well as by measuring the ova diameter. Gonadosomatic index of the species has also been widely used to indicate the maturity and periodicity of spawning and predicting the breeding season of the fish, and can be calculated by the following formula:

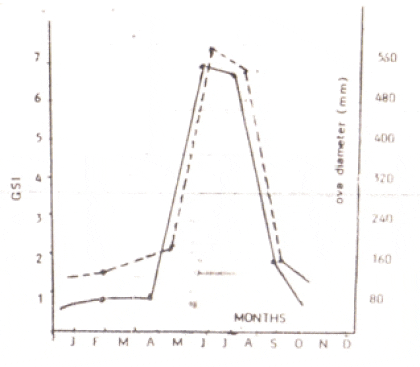
Gonadosomatic index (GSI) of the fish increases with the maturation of the fish, being maximum during the peak period of maturity and declining abruptly after spawning. The GSI of river catfish (Glyptothorax pectinopterus) was found to be highest in June and least in November (Khanna and Pant, 1967) which coincides with the ova diameter as shown in Fig.11 (below). The breeding season of this fish extends from June to July.
Studies on the GSI and histology of the ovary in snow carp (Schizothorax plagiostomus) show that this fish spawns twice in a year (Shrestha and Khanna, 1979). In this species the first spawning occurs from mid-September to October. During winter, the fish carries mature as well as immature oocytes in dormant phase, and second spawning takes place in March on the return of favourable conditions of temperature, rainfall, pH etc.
Seasonal changes in the ovaries of teleosts have been studied histologically as well as by measuring ova diameter, and with help of gonadosomatic index.
- Spawning takes place between March and early June in freshwater catfish (Mystus seenghala) (Sathyanesan, 1962),
- From April to September in Tor tor (Rai, 1957),
- From June to September in Clarias batrachus (Lehri, 1968),
- From July to September in Channa punctatus (Belsare, 1962),
- From June to August Channa gachua (Khanna and Sanwal, 1971),
- From April to August in Rasbora daniconius (Raizada, 1971), and
- From October to December in Suchizotharax richardsonii (Bisht, 1972).
The length of the breeding period is thus extremely variable, and some species spawn only once, others twice, while still others may spawn several times during a year. Both photoperiod and temperature are considered important factors controlling the maturation of gonads in fishes (Hazard and Eddy, 1951). Variations in the duration of light and/or temperature, may influence the rate of gonadal development. Bullough (1940) has reported the existence of an internal rhythm in Phoxinus.
Atretic oocytes (Pre-ovulation corpora lutea)
All the developing oocytes of a fish ovary do not reach maturity and undergo degeneration and resorption. These are known as the atretic oocytes and are conspicuously seen during pre-spawning immediately after spawning.
The process of atresia has been described by several authors in Indian fishes like Mystus seenghala and Walago attu (Dixit, 1956), M seenghala (Sathyanesan, 1961), Channa punctatus (Belsare, 1962), Tor tor (Rai, 1966), Clarias batrachus (Lehri, 1967), Glyptothorax pectinopterus (Pant, 1969), Channa gachua (Khanna and Sanwal, 1972).
The oocytes are surrounded by follicular cells that from a syncytium. In some cases these follicular cells are enable to produce healthy, mature ova, which become degenerate and disappear. The follicular cells may undergo hypertrophy, finally developing into an enlarged glandular mass, called the pre-ovulation corpus luteum. Details of the process have been described by Bretschneide and Duyvene d Witt, (1947), and Hoar (1965). During the recent years two processes for the formation of atretic eggs have been described such as (a) hypertrophic atresia, and (b) non-hypertrophic atresia.
Hypertrophic or phagocytic atresia
In the early stages, the nucleus of the oocyte disappears, and there is erosion of the vitelline membrane (zona radiata), which become wavy, losses contact with the granulose cells, and may become broken at some points.
The follicular cells which from a syncytium in the healthy oocyte, acquire cell wall, and undergo hypertrophy. There is liquefaction of the yolk and in advanced stages of the atresia, the follicular cells increase both in size and number, and invade the cytoplasm of the oocyte engulfing the cosinophilic yolk.
The atrectic follicle is richly vascular. The zona radiata becomes broken and disappears, while the granulose cells become vacuolar and digest the yolk. Ultimately, the area occupied by the oocyte, is filled with a loose mass of follicular cells which possess yellow or orange granules. As the yolk is consumed, the follicular cells collapse, shrink and disappear. The follicular cells generally have indefinite cell outgrowth and digest the yolk by active phagocytosis. Young oocytes also undergo atresia, but this is rare. In a few oocytes, the hypertrophy of the follicular cells is confirmed to one point, so that a structure referred to as "cell pearl" is formed.
Non-hypertrophic atresia
The nucleus of the oocyte disappears and the vitelline membrane is eroded. It becomes folded at one or more points and may even break. There is no hypertrophy of the granulose cells. The yolk becomes basophilic and finely granular, and is largely removed through an opening. The vitelline membrane finally collapses, and the cells from a small ball which disappears later.
The cause of significance of the atresia is not fully known. It has been suggested that the atresia is due to low gonadotropin content of the pituitary, which is not able to maintain healthy oocytes (Rai, 1966). As these bodies are formed from the follicular cells as in mammals, they are considered homologous with the corpus luteum of mammals.
The pre-ovulatory corpora lutea or corpora atretica may or may not produce hormones, but it is an active organ, and its formation is related with changes in the pituitary gland (Hoar, 1957). Beach (1959) has concluded that the corpus luteum-like structures in fishes, are associated with the removal of yolk rather than the secretion of the hormones.
Many investigators are of the view that these structures do not correspond to corpora lutea of mammalian ovary. Recently Chieffi (1961, 62, 67) has produced histochemical evidence in support of an endocrine function of the corpora lutea in elasmobranches.
The discharged follicle (post-ovulatory corpus luteum)
Besides the pre-ovulation corpus luteum (corpora atretica) described above, comparable structures develop from the follicula cells after the discharge of the mature ovum.
After ovulation, the theca and the follicular cells are left behind and appear as scars in the ovary. After the discharge of the egg, the remnant of the follicle shows broken walls, which later becomes continuous. The cells of the granulose layer proliferate and enter into the lumen of the follicle so as to fill all available space.
These structures are called post ovulatory corpora lutea, and are ultimately reabsorbed by phagocytosis. These structures have been compared with the corpus luteum of mammals, because they develop similarly from the follicula epithelium and are believed to secrete hormones. However, there is little evidence to show their secretory nature and endocrine function. The use of the term "corpus luteum" for these bodies has, therefore been questioned by several authors.
Length weight relationship and condition factor
Several studies have shown that the length of the fish maintains a steady relationship with the weight ie, an increase in length is related with the increase in weight of the fish, but the degree of this relationship varies with the species of the fish and there is no effect on season or sex. If the numerical volumes of the average weight are plotted against those of the average length, a parabolic curve is obtained. If the average log length is plotted against the log weight, a straight line is observed. For such a study, total length and weight of a large number of fish are recorded in fresh condition, and the average values are analysed separately in groups of fishes consisting of individuals varying by 5mm, and the mean values are then calculated.
Based on the data obtained as above, the condition factor is calculated by using the formula: K = W / w
Where, K = relative condition factor
W = observed weight of the fish
w = calculated weight of the fish
The values of condition factor are useful in fishery investigations, and can be used to express the relative health or robustness of the fishes. The values of K differ with the season and are influenced by maturity of gonads and spawning. The condition factor is maximum during the spawning season.
Ovulation and fertilisation
The egg gradually enlarges and projects into the lumen of the ovary. At ovulation the follicle ruptures and the egg escapes from the aperture through amoeboid movement. The forces causing the rupture are not known and the muscle fibres present in the ovarian wall may be responsible for it. In teleosts having cystoarian ovary, the ova are discharged into the oviduct, while in those fishes which have a gymnoarian ovary, the eggs are discharged into the body cavity. The muscles of the oviduct and those of the body wall contract causing movement of the ova through the oviduct.
Fertilisation is external in water in majority of the teleosts and internal fertilisation also takes place in some species. Spermatozoa and eggs are discharged into water in close proximity. The sperms become very active in water, and survive only a few minutes, during which the fertilisation takes place which may be controlled by several chemicals released by both sperms and eggs.
In some species of teleosts, fertilisation is internal and the urogenital papilla or anal fin is enlarged or modified for the transfer of sperms. In cyprinodont fishes like Gambusia, the anal fin rays of male are modified to form an elongated copulatory organ which is tubular or grooved and is called the gonopodium. In Scorpaenidae and Embiotocidae, which include viviparous teleosts, there is no gonopodium and the sperms are transferred by genital papillae of the male coming into contact with the cloaca of the female.
In Cottidae, the genital papillae are enlarged to from the penis in the male. Wiesel (1949) has described erection tissue in the genital papillae of both males and females. In Apogon imberbis, the female has elongated genital papillae, which is introduced in the male for receiving the sperms (Garnaud, 1950). Female Rhodeus has a much elongated genital papilla, to transfer the eggs into the shell of bivalve Anodonta where they are fertilised by the sperm shed in water and develop inside the shell.
In the viviparous teleosts, fertilisation generally takes place while egg is within the follicle (follicular gestation) or development takes place within the ovarian cavity (ovarian gestation).
Eggs
The number of eggs produced by a single female differs considerably and depends upon several factors like her age, size, condition and species. The egg is generally surrounded by a shell but when it leaves the ovary, it is enclosed in a vitelline membrane. Generally, the egg is spherical or oval in shape and has some amount of yolk in it.
Eggs of bony fishes are of two main types. Pelagic eggs are buoyant and provided with a thin, non-adhesive membrane, while demersal eggs are heavy and sink to the bottom, and are covered by a hard adhesive membrane. Sticky, demersal eggs become attached to the debris of the bottom and are prevented from being swept away along the current of water at the time of deposition.
Marine fishes produce either pelagic or demersal eggs but the eggs of fresh water fishes are generally demersal. Pelagic eggs are of small size and a single large oil globule may be present on the surface of its yolk.
The eggs of some species (Scomberisocidae, Belonidae and Exocoetidae) have sticky threads for attachment with some object or with each other.
Fecundity
Fecundity can be defined as "the number of ova that are likely to be laid by a fish during the spawning season". The number of eggs produced by a fish differs in different species, and depends on the size and age of the fish. It may also differ in different races of the same species. Thus, fecundity is a measure of the reproductive capacity of a female fish, and is an adaptation to various conditions of the environment. It can be estimated by any of the following methods:
- In the volumetric method, the total volume of the ovary is measured. Small pieces of the ovary are taken in random samples from the anterior middle and posterior parts of the ovary, and the number of ova in each sample is counted along with the volume of the sample. The total number of ova in the total volume of the ovary is then calculated.
- In the gravimetric method, the formalin preserved ovaries are used. After determining the weight of the ovary, three small samples of 100 mg each are taken at random from the anterior middle and posterior part. The number of ova in each sample is counted under a binocular microscope. Total number of ova are then calculated as: F = S x OW / 100.
Here, F = Fecundity
S = Average number of ova obtained from three samples of 100 mg each
OW = Total weight of the ovary
The fecundity of a number of species has been studied by several investigators. Fecundity of Cirrhina mrigala varies from 75,900 to 11, 23,200 when the length ranges from 349 – 810mm in length (Hanumantha Rao, 1974). In Labeo gonius ranging between 270 – 490mm, the fecundity varies from 47,168 to 3,80,714 (Joshi and Khanna, 1981), but according to Chondar (1970) the values are 2,73,955 to 5,39,168 in the fish ranging from 270 – 490mm in length. The differences in the values are possibly due to the collection from different habitats.
Fecundity of the fish is closely related to the fish length and weight. A straight line relationship has been observed between the fecundity and the fish length, and the fecundity and fish weight in a number of species.
Knowledge of the fecundity of fish is useful in fishery management and the information regarding the possible number of eggs and fry likely to be produced can be used for selecting the fish of high productivity for fish culture.
Sexual dimorphism, courtship and parental care
Some species of fishes exhibit well marked sexual dimorphism. In the Chondrichthyes, the eggs are fertilised within the body of the female and the males are provided with claspers or myxopterygia for transferring the sperms into the body of the female. Among teleosts, an intromittent copulatory organ is present only in those species like Mystus seenghala in which a conical genital papilla is present and fertilisation is external.
In Cyprinodonts (eg Gambusia) the vas deferens is produced into a tube up to the end to the interior rays of the anal fin. In the Poecillidae also the males are provided with complicated intromittent organ developed from modified anal fin rays, ending in curved hooks, spines and barbs. The male four-eyed fish, Anableps, is also provided with a special tube for copulation. In all these fishes, the fertilised ova are retained within the body of the female and young ones are born.
In several species of fishes, the sexual differences are not related with copulation between male and female, and are generally well marked during spawning season. In most of the teleosts, the females are larger in size with enlarged rounded belly during the breeding season. A common secondary sexual character is the brighter colour of the body and fins in the male as in Cyprinodontidae, Cichilidae, Labyrinthidae and Labridae.
In a number of Cyprinids, the male becomes more brighter in colour during the breeding season. The male Bow-fin (Amia) has a characteristic black spot at the base of the caudal fin. Some species show differences in the shape of their fins and the rays may be prolonged to from long feelers in the male. In some cat fishes of the family Loricaridae, the sexes differ in the shape of their snout, mouth, lips and in the development of bristles on their head. In the male sword-fish (Xiphophorus) the lower lobe of the caudal fin is drawn out to form a long blade like structure. The male of many cyprinids develop horny tubercles on the head and body, especially during breeding season. In some Gobies and Blenies, the male possess enlarged canines.
The courtship of the female by the male consists in swimming merry go round fashion in her vicinity during which the male exhibits his brilliant colours, this is beautifully exhibited by some species as the Siamese much excited during the breeding season, that they fight with other males inflicting considerable damage to one another. The males of certain Gobies also fight with each other and the victorious one shows its bright colours to the female.
Many fishes do not care for their eggs and leave the spawning grounds after fertilisation. But some species have evolved various methods to ensure proper development of the eggs which may be protected by one or both the sexes. Some fishes prepare crude nests for egg lying. At first a suitable place for preparing the nest is selected and some species defend the place till death.
Males of many species like the Darters (Etheostoma) sun fishes and the Cichilids, prepare a shallow basin-like nest and all stones and pebbles are carefully removed from its bottom. The eggs are laid in the nest and the male after fertilising them keeps guard over them till the young ones are hatched.
A few species, however, leave the nests unprotected. Some fresh water fishes prepare a nest by clearing space among aquatic vegetation. The African lung fish (Protopterus) prepares a simple nest in the form of deep hole in swampy places along the river banks. The male prepares the nest and after spawning keeps guard over it, occasionally aerating the water by his slow body movements. The South American lung fish Lepidosiren, also prepares a nest in the form of a burrow in swampy places and the male develops highly vascularised filaments on the pelvic fins, which probably serve to secrete oxygen into the surrounding water. The male Bowfin (Amia) prepares a crude circular nest among aquatic vegetation. The fertilised eggs are then protected by the male who keeps guard over the nest till, the young ones are hatched. The young ones are allowed to leave the nest in a body under the protection of the father. Both the male and the female of some cat fishes of North America (Amiuridae) prepare a crude nest in the mud for egg lying. The nest is sometimes provided with a protective cover of logs, stones etc.
The male Gasterosteus aculeatus, prepares an elaborate nest before courtship is begun. He selects a suitable place among the aquatic plants where water flows regularly but not swiftly. He then collects plant material which is pressed into a clear area of the nest. A sticky substance is produced by the kidneys of the male and serves to join the plant pieces together. When sufficient amount of the plant material is collected, the male burrows through its centre, thus making a small tunnel, through which he brings a ripe female for egg lying. After the eggs have been fertilised, the female leaves the eggs and the male keeps guard over it. In the early stages of the development, the male "fans" the developing embryos by sending a current water through the nest. Later, the male stops the fanning activities and keeps close watch over the brood, not allowing any young one to go astray.
Many of the Labyrinth fishes prepare a nest by blowing bubbles of air and sticky mucus. These adhere forming a floating mass of foam on the surface of water. The male fighting fish, Betta, also prepare a nest in the same way. The fertilised eggs are collected by the male in his mouth who gives them a coating of mucus and sticks them to the lower surface of the foamy nest. The nest is then protected by the male. The male paradise fish Macropodus, also prepare a similar foamy nest but in this species the eggs are lighter than water, and rise to the nest without the active participation of the father.
The European Bitterling, Rhodeus, takes extra care for the protection of eggs. When the female is ready to spawn, the oviduct extends out to form a long tube acting as an ovipositor which is used to deposit the eggs within the valves of the fresh water mussel. The male fertilises the eggs as they are laid. After hatching, the fry leave the host and thus remain well protected from the enemies. In some species, eggs develop within the mouth of the parents. In many Cichilids, the female carries the eggs in her mouth. After hatching also, the young fry do not leave the shelter for some time and swim about in water very near the mouth, so that they can return to it in case of danger. In the cat fish, Aureus, the male carries the eggs and young one in his mouth, who does not take food during this period.
The Butter fish (Pholis) rolls the eggs into a rounded ball and then one of them remains on guard by coiling round it. It is not definitely known whether the male or the female or the both take part in protecting the eggs in this way. A cat fish, Platystacus, a cat fish of Brazil exhibits an interesting phenomenon of parental care. The skin at the lower surface female becomes soft and spongy and it presses the body against the eggs so that each one of them get attached to the skin by a small, stalked cup. The eggs remain fixed in this position till hatching.
In the Pipe fish (Syngnathus) and the sea horse (Hippocampus) the fertilised eggs are transferred by the female into the brood pouch of the male who carries them till the time of hatching. Finally, highest degree of parental care is exhibited by those species which are viviparous and produce young one. Among the teleosts, this is seen in Cyprinodontidae (Lebistes, Gambusia) Poecilidae, Anablepidae, Jenysiidae, Hemiramphidae etc.
Migration for spawning
Many marine fishes migrate to the rivers for spawning. These are called anadromous fishes and include Lamprey (Petromyzon), Salmon and hilsa. Most of these fishes produce a very large number of eggs that are left uncared for. The Atlantic Salmon (Salmo salar) spawns during November and December. They ascend the rivers during the breeding season and on entering the fresh water give up feeding and lose weight. The silvery colour is replaced by a dull reddish brown shed. Skin becomes spotted with red, orange and large black spots. Suitable spawning grounds are selected and the fish segregate in pairs. Shallow saucer-like nests are prepared where spawning takes place. After spawning, the return journey to the sea is undertaken, but very few male survive to breed a second time. Females that are able to reach the sea, start feeding and soon recover their normal condition and silvery colour. Salmon does not usually spawn more than three times in its life span of eight or nine years. The king salmon (Onchorchus) also has a similar breeding habit.
The sea-lamprey (Petromyzon) is also anadromous in habit and ascends the rivers for spawning, during spring or early summer months. Lampreys also undergo considerable changes in colour during breeding time, and the males and females show marked differences in appearance. They reach clear, shallow, fast flowing streams, provided with sandy bottom and scattered stone pieces. A crude nest is prepared by clearing a circular or oval space, 2-3 feet in diameter. The stones are moved and piled up at the lower end of the nests. Usually the male reach the suitable place earlier, and starts nest building. Each male is later assisted by a female. Some times a second female also joins the pair. They attached themselves to stones by their suctorial mouths, and drag them to the lower end of the nest. Mating takes place under the ready nest. The females attaches her to a large stone near the upper end of the nest, the males seizes her by the top of the head and coils round her body. They vibrate the hinder part of their body vigorously; extradite the ova and sperms simultaneously. The eggs are coiled by a sticky substance and sand. The male and female now separate and start removing the stones from above the nest, so that more sand is carried by the water to cover the eggs. The pair leaves the nest after all the eggs have been covered. By this time they are very much exhausted and fall an easy prey to the enemies and do not survive.
The common fresh water eel, Anguilla anguilla (European eel) also undertakes long journeys to reach the spawning grounds, but this migration is from rivers to the sea (catadromous). Eels are are of two types
- Yellow eels: Represent their feeding and growing phase.
- Silver eels: Represent their breeding phase
Yellow eels are found in both salt and fresh water and vary in length from a few inches to several feet. With the approach of autumn, a number of yellow eels become silvery and prepare the migration towards spawning grounds. The migratory male fish belongs to the age group 8 to 10 years, while the females are 10 to 18 years old. At this stage they stop feeding, eyes become large, lips thinner, the pectoral fins more pointed and darker in colour. The colour of the body becomes silvery on the sides, with blackish back. These characters become more pronounced with the approach of breeding time. The reproductive organs develop and the alimentary canal becomes shrunken. The eels migrate down the rivers, reaching the sea in the late summer. It is believed that these eels cross the Atlantic Ocean, covering three to four thousand miles, and ultimately reach their spawning grounds in the Western Atlantic and south of Barmuda. It is assumed that the eels spawn at a depth of about 400 meters below the surface at 16 to 17 °C. It is likely that both the spawners die after spawning.
The eggs float for sometime and hatch at a depth of 60-150 feet. The larvae are called the Leptocephali, and are flat, leaf-like, tiny creatures, provided with long, needle like teeth for feeding. To start long homeward journey they move in the easterly direction, grow rapidly during the first few months, and when they reach the coasts of Western Europe, they are about three inches long and two years old. At this stage they undergo metamorphosis, during which time they stop feeding, lost teeth, and the body assumes a cylindrical shape. They are transparent and are called elvers or glass eel which acquires new teeth. The elvers, when become three years old, start ascending the rivers in huge numbers, and are able to cross all obstacles and finally on reaching their suitable resting place they feed and grow for some years and ultimately develop into yellow eels. Finally on attaining maturity, they finally change into silver eels, and start for their own breeding migration to the sea, which ends in death.
Another closely allied species is called the American eel, Anguilla rostrata, and is found in the rivers and along the coast of North America. The breeding area of both the species overlaps, and the larvae of both the species are found together. The larvae of the American eel migrate towards the rivers of North America. According to Schmidt, the larvae of the American eel grow more rapidly and become metamorphosed into elvers in one year, while the larvae of the European eel take three years to become elvers. The distance to be covered by individual species is dependent on larval lifespan. The long larval life of the European eel is an adaptation for covering a great distance to approach breeding grounds which is located far away from the coasts.
However, a British Zoologist, Dr Tucker, has put forward another hypothesis. He opined that the two species of eels are really one and the same. The small morphological differences between the two are due to environmental factors. According to Dr Tucker, the European eels are not able to survive the presumed 3500 miles journey from Europe to the Sargasso Sea, and they do not return to the spawning grounds as also mature European eels have not been observed in the said area. The American eels are able to reach the spawning area as they cover shorter distance. Thus it appears that the European eels are also derived from the American eels. The actual spawning area determines whether the larvae will be carried towards America or Europe, along with the currents. The temperature and other ecological factors determine the larval age. Extensive field and experimental studies needs to carried out before drawing conclusion.
Development
Cleavage and the formation of blastula
The development of an egg begins soon after it is fertilised by a sperm. The egg of bony fishes has a relatively large amount of yolk, which remain segregated from the active superficial cytoplasm. Cleavage is confirmed to the superficial layer of the cytoplasm and is incomplete underside (meroblastic). In the earlier stages cleavage planes are all vertical so that all the blastomeres lie in one plane only. The blastomeres are separated from each other by furrows but lie over the yolk. In the later stages, cleavages occur in the horizontal plane also, so that the blastomers become arranged in more than one row. The marginal cells are in contact with the yolk. The disc of cells thus formed on the animal pole of the egg, is called the blastoderm. The central cells of the blastoderm divide to form a number of "free" blastoderms which subsequently arranged on the top of the yolk so as to form a layer of cells called the periblast. The space between the blastoderm and periblast is the blastocoel and the embryo now is in the blastula stage. The blastoderm gives rise to embryo proper while the cells of the periblast probably serve to digest the yolk, and supply it to the developing embryo.
Fate map of blatula
It is possible to identify various regions of the blastula wall that are destined to give rise to specific organs in the embryo. Thus a fate map of the teleostean blastula can be constructed showing the presumptive ectoderm, endoderm, mesoderm, notochord, neural plate etc. In the fish blastula, the areas which are destined to give rise to the organs of the dorsal region of the animal, tends to be concentrated towards one side of the blastodisc. This indicates the posterior end of the future embryo. At this end and along the margin of the blastoderm, lies the presumptive endoderm which gives rise to gut. Anterior to this lies the presumptive notochord, and the centre of the blastoderm, holds the area of the nervous system. The presumptive mesoderm lies along the sides of the areas destined for endoderm, notochord and neural plate. The mesodermal area extends mainly along the margin of the blastoderm and in Fundulus, is not continuous at the anterior end due to the presence of presumptive epidermis. But in Salmo the presumptive mesodermal area is present all round the margin of the blastoderm.
Offspring numbers and parental care
Fish may lay eggs, ie be oviparous, or give birth to live young. Those species which give birth to live young can be further divided into the truly viviparous species, in which the developing embryos are supplied with nutrients by the mother, and ovoviviparous species, in which there is egg retention but no additional nutrient supply from the mother.
Within this broad framework of the three basic modes of reproduction- oviparity, ovoviviparity and viviparity- there are large interspecific differences with respect to both the numbers of offspring produced, and the degree of protection and care given to the developing eggs and young by the parents. Within the oviparous species, for example, the variations may range from the production of large numbers of freely floating pelagic eggs at one extreme to nest building and extensive parental care at the other. For example, many marine species produce large numbers of small pelagic eggs, whereas stickleback species (Gasterosteidae) build nests and engage in elaborate courtship and parental behaviours.
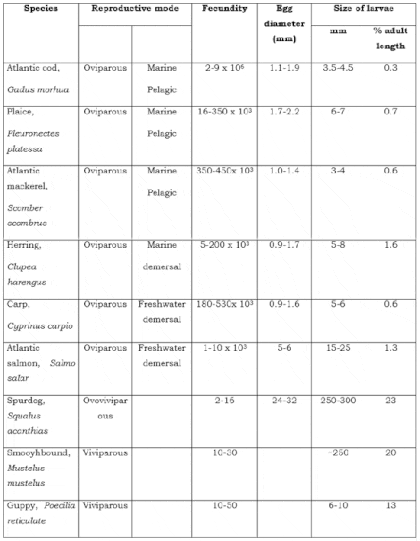
By studying the relationships between fecundity (the number of eggs produced per female), egg size and degree of parental care it is possible to make some generalisations about reproductive patterns in fish (Tables 9.1 and 9.2). Thus, it can be said that the following general principles appear to apply.
- Fecundity tends to be high where eggs and sperm are liberated freely into the water, is less in those species giving some form of protection to the eggs and is lowest in species that show parental care.
- Fecundity and egg size are inversely related, that is fish produce many small eggs or a few large ones.
- The production of pelagic eggs is largely restricted to marine species. Freshwater species usually have non-buoyant, sticky eggs that are deposited on the substrate, or they build some type of nest.
- Fecundity is higher in oviparous than in either ovoviviparous or viviparous species.
These generalisations refer to interspecific differences, but even within species there can be considerable variations in both fecundity and egg size (Table 9.1). There may, for example, be considerable intraspecific variability in egg numbers, both with respect to differences between females spawning within a given season, and with respect to the fecundity of a given female in consecutive years or breeding seasons. Variations in fecundity of up to 45 percent have been recorded for plaice, Pleuronectes platessa, and variations of about 25 percent, 34 percent and 56 percent have been found for pike, Esox lucius, hering, Clupea haregus, and haddock, Melanogrammus aeglefinus, respectively.
The intraspecific variations in egg size and fecundity can usually be traced either to differences in the sizes, or ages, of the females making up the spawning stock, or to year-to-year fluctuations in the food supply. For example, in many species of fish there is a clear relationship between fecundity and size of the female. Larger females tending to produce more eggs than do smaller conspecifics.
Intraspecific variations in fecundity and egg size may also be related to the time of spawning. For example, it is frequently reported that fish spawning late in the season tend to produce smaller eggs than do early spawners.