This study attempted to address the role of each mode of nutrition on the developmental growth and survival from larval, to post metamorphic microscopic clams to visible 2-3 month old juveniles. Studies on the different levels of the algae and dissolved inorganic nitrogen (DIN) were conducted to determine optimum rates to supplement the heterotrophic and autotrophic modes of feeding of the clam-symbiont association, respectively. This study has practical importance in the culture rearing protocol of giant clams in Aqaba, Red Sea.
Experimental set-up
Four treatments were implemented as follows: Algal-feeding (A), Nitrogen-spiking (N), Combined Algae + Nitrogen (NA) and Control (C). Uptake rates for both DIN as ammonium and culture microalgae are monitored at particular life stages as follow.
Two experimental trials were done using Tridacna maxima clams. In the first run, 4-day old larval clams were collected from T. maxima cohort spawned shortly stocked at 3 veligers/ml. Clams were placed in 1.5L bins with separate inlets and drains. Bins were arranged in a block and placed in one outdoor tank in a water bath supplied with 5mM filtered seawater.
There were 4 replicate bins per treatment. Nutritional conditioning commenced by daily feeding of dissolved inorganic nitrogen in the afternoons, in static water to allow clams access to the supplements until morning when flow commenced. Approximately, 20mM ammonium chloride was spiked in the bins.
Based on the results in the first run, modifications were made in the second run to check the uptake due to clams alone. For each treatment, 2 replicate-bins were stocked with clams and the other 2 without clams as controls. Higher stocking density was also used at 10 villagers/ml to see the effect of density on the nitrogen. 3-day old larvae were collected from T. maxima cohort_ spawned recently. Clams were still subjected to similar nutritional conditioning as in first run.
Water samples were taken at 4-hour intervals from feeding/spiking (i.e., 0, 4, 8, 12 hours), aliquot samples of 20 ml were collected in each bin, filtered through a 5mM sieve, for both ammonium measurements. Ammonium analysis was implemented using spectrophotometer following Strickland and Parson (1972).
Uptake of nitrogen ammonium
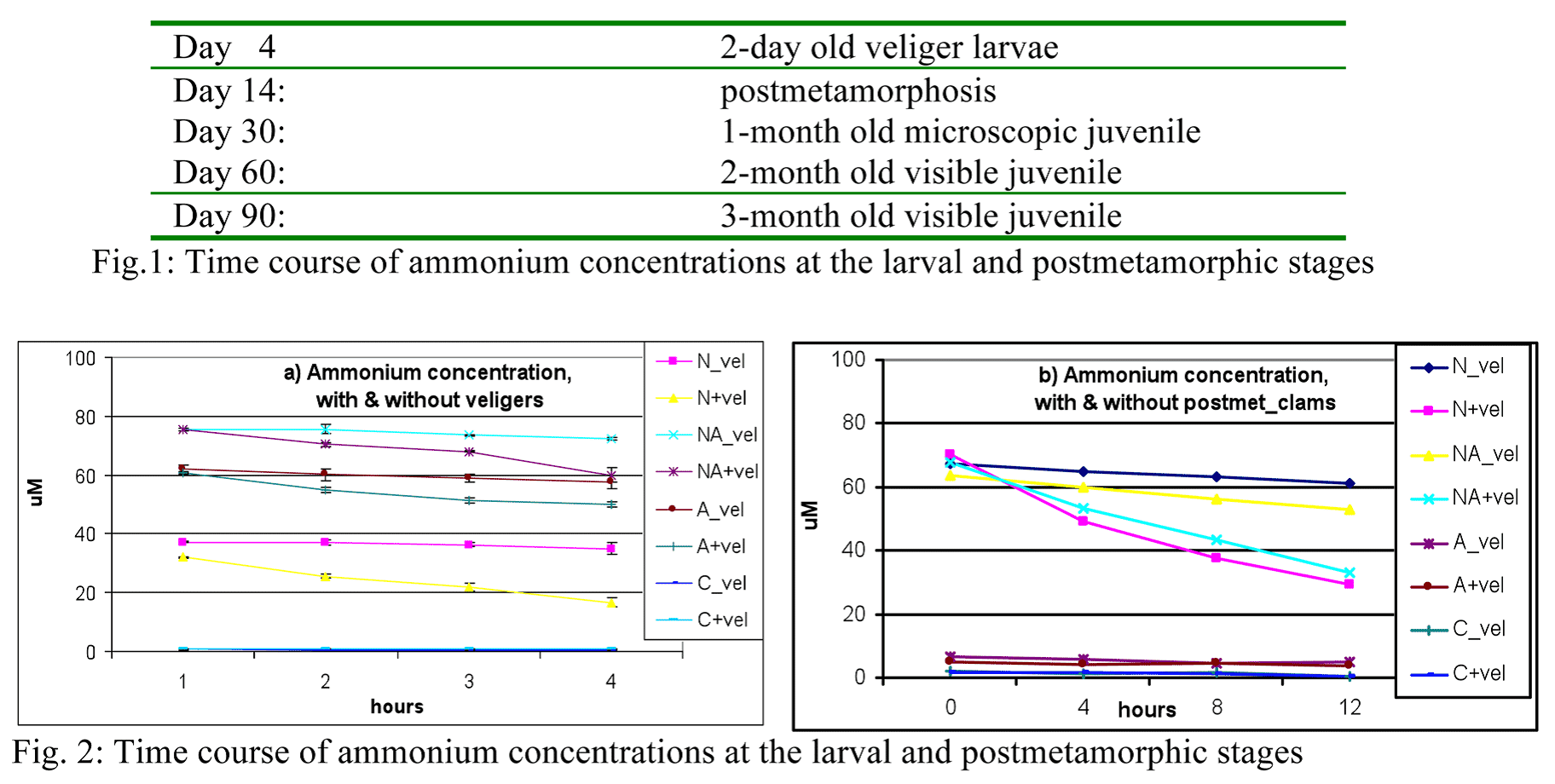
Figures 1a-b shows the ammonium concentration in all treatments at the larval and post metamorphic stages, respectively.
In both figures, there is marked difference in the ammonium concentration between the spiked treatments (N and NA) ranging from 2.37-6.27 mM and non-spiked treatments (A and C) at 0.22-0.62 mM. A continuous decrease of the nitrogen level was observed in N and NA over time, compared to relatively same concentration over time in A and C.
The uptake rates at 4-hour intervals in both stages were derived from difference of concentration from t1-t0 and shown in Table 1. It was observed that in both veligers and postmetamorphosis, there was a fast uptake in the first 4 hours upon spiking for all spiked treatments as shown in Fig. 2. In the succeeding hours, uptake rates decreased and ranged was similar for both stages.
Was this decrease in ammonium level solely due to the uptake of the clams alone or other factors? Figs. 2a-b showed the ammonium levels of the treatments with clams and without clams at larval and postmetamorphic stages, respectively. In the veligers, the spiked bins without clams showed relatively same ammonium concentration over time, from 37.1 to 35.0 mM for N and 75.5- 72.3 mM for NA. In comparison to the bins with clams, the concentrations were lower and there was a continuous decrease of ammonium over time, 32.0-16.7 mM for N and 75.3-59.9 mM for NA. With the postmetamorphosis
clams, the same trend was observed, where the ammonium concentrations were lower and decreasing for the bins with clams.
(Note in Fig. 2a, the algal-fed (A) treatment showed high ammonium concentration at veliger stage, probably due to some remaining nitrogen from the fertilizer not fully utilized by the algae yet. Nevertheless, the bins with clams showed lower and decreasing nitrogen compared to the bins without clams, showing similar trend as mentioned above.
To get the actual uptake of ammonium by the clams alone in the spiked treatments, the average concentration (at 4-hour intervals) in the bins without clams was subtracted from the bins with clams to get the difference, which was due to clams alone. The resulting differences showed a faster uptake within 4 hours upon spiking in N and NA treatments, similar for both veligers and postmetamorphosis clams (see Fig. 2). Uptake still continued over time but at lower rates for all treatments in both stages (except NA_veligers). In the both runs, the postmetamorphic clams were observed with a higher
consumption of ammonium at the first 4 hours compared to the veligers. In the succeeding times, no trend was observed between the two stages. In the first run where stocking density was at 3 veligers/ml, ammonium uptake of the veligers ranged from 0.09-0.54 mM/hr compared to higher density (10 vels./ml) in the second run which ranged from 0.13-1.71 mM/ hr. For the post-metamorphic clams, lower clam density showed uptake range of 0.05-1.16 mM/hr compared to the higher density of 1.52-4.65 mM/hr. These differences in ranges at both stages showed the uptake of ammonium was influenced by the stocking density of the clams.
October 2013



