Local and global environmental impacts of tourism are intensifying due to rapidly expanding human populations. Marine shorelines are particularly vulnerable because of the massive residential shift toward coastlines and the fact that seashores remain among the most alluring tourist destinations . Tourists and single-day visitors sensu affect beaches in diverse direct ways, including trampling, vehicle use, camping, shellfish harvesting, beach grooming, and many other factors. However, long-term studies measuring human impacts on shoreline habitats are scant and fragmentary. In particular, the removal of dead shell remains by tourists represents one of the most understudied and least understood processes associated with human activities along marine shorelines.
Rigorous assessments of shell removal by tourists are needed because skeletal materials left behind by dead organisms perform many important environmental and ecosystem services. In pristine coastal habitats shells can occur in great abundance and serve multiple functions, from beach stabilization to building materials for bird nests. Also, shells may be intermittently submerged or transported into subtidal settings where they can provide various ecosystem services, including shelter for diverse endobiolithic algae, substrate for seagrass meadows, colonization sites for encrusting organisms, or armored protection for hermit crabs and fish. Shell material is also continuously dissolved in most coastal areas resulting in elemental recycling back into the global marine reservoir. The tourism-related removal of shell material from shorelines may thus have diverse environmental consequences.
Diverse processes can induce seasonal or decadal changes in shell abundance along shorelines and multiple hypotheses can be postulated a priori to explain variation in shell abundance through time. First, changes in local hydrodynamics may alter the magnitude of onshore transport of shell material. This model predicts a significant correlation between proxy measures of onshore transport (e.g., wave energy) and shell abundance. Second, changes in ecosystem structure and population dynamics (especially spawning and mortality patterns) may induce variation in local shell accumulation rates, which may be partly controlled by input of newly dead specimens. This model predicts seasonal or decadal-scale correlations between temporal ecosystem changes and trends in shell abundance and also implies possible changes in rank abundance of dominant species or other ecological proxies (e.g., frequency of predation events and perspecies size-frequency distributions). Finally, as postulated here, changes in tourism activities and tourism-related beach management practices may be responsible for changes in shell abundance.
This last model predicts an inverse seasonal (or decadal) correlation between shell abundance and tourist arrivals.
In this study, local changes in shell abundance observed over the last 30 years were evaluated for one Mediterranean beach using two quantitative datasets from the same embayment, one collected more than 30 years ago and one collected in recent years. During a time interval of three years (7/1978 - 7/1981), quantitative surveys of shell material were conducted monthly along Llarga Beach Salou, Catalonia. The same shoreline transects were resurveyed 30 years later (2008–2010). Because the sampled beach has not been altered considerably over the last 30 years, the resulting comparative data should be particularly suitable for testing the hypothesis that an increase in local tourism leads to the accelerated shell removal. Specifically, we evaluated the three models outlined above regarding changes in shell abundance in a local shoreline habitat. We tested for correlatives between the intensity of local tourism and shell abundance at two temporal scales: intra-annual/seasonal and multi-decadal. Within the same reference timeframe, we evaluated putative roles of other causative processes unrelated to tourism that could have led to shifts in shell abundance, either seasonally or through time, and explored global implications of the tourism-related removal of shells from coastal environments.
Our focus on removal of empty shells due to tourism in an area that has remained relatively unchanged differs in goals from studies targeting regions that experienced substantial ecosystem degradation through time. Also, previous projects aimed at assessing the impact of human activities on live shellfish, including subsistence and recreational harvesting, collecting for curio trade, bait collecting, or inadvertent trampling of live organisms. Instead, we focused here on activities that may be intuitively perceived as least harmful: the impact of beachcombing (leisurely collecting, inadvertent trampling, use of recreational vehicles, etc.) on empty seashells scattered along marine shorelines.
Results
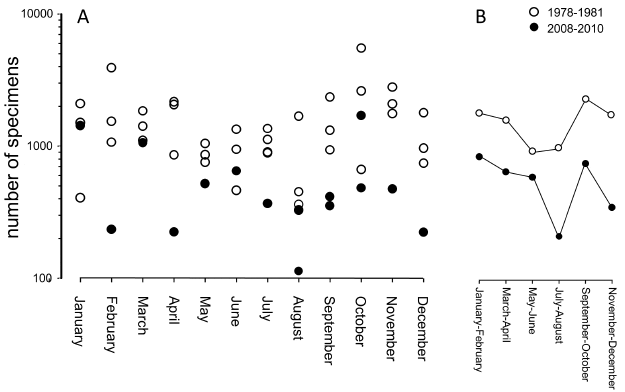
A comparison of 1978–1981 and 2008–2010 surveys indicated that shells at the shoreline of Llarga Beach (Fig. 3) were almost three times more abundant three decades ago: on average, 1506.5 specimens were recovered per transect in 1978–1981 compared to only 578.3 specimens in 2008–2010. The decline in shell abundance was comparable in magnitude for the tourist season (July–August) (70.1 per cent) and for other months (60.0 per cent). When 1978– 1981 and 2008–2010 transect estimates were binned bimonthly (due to small number of transects for some of the winter months, monthly bins were inadequate), there was a remarkable congruence in seasonal changes in shell abundance with notable declines in shell quantity observed in the late spring, summer, and the late fall (Fig. 3B). One notable difference between the two timeintervals was a much steeper decline in average specimen abundance, which was observed only for the July-August months for the 2008–2010 sampling interval (Fig. 3B).
The maximum monthly wave energy [MMWE] for 1978–1981 and 2008–2010 was not correlated significantly with shell abundance [SA] (Spearman r= 0.10, p= 0.46) and the pattern persisted for first differences (month-to-month changes) [ΔSA] (r =20.18, p= 0.21). When MMWE values were downweighted by the length of a time lag between the last day in which wave height was at monthly maximum and the day of sampling [MMWE*=MMWE/#days], correlations remained non-significant: (SA: r = 0.26, p =0.06 and [ΔSA]: r=20.23, p =0.10, respectively). All correlation coefficients were low (r,0.3), the first differences [ΔSA] yielded correlation coefficients with an opposite sign relative to the raw correlations [SA], and none of the tests was significant. When data were subdivided by season, time-interval, or both, correlation coefficients either remained low or were noninterpretable due to excessively small sample sizes.
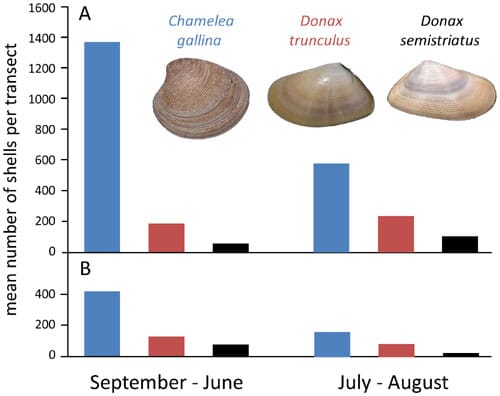
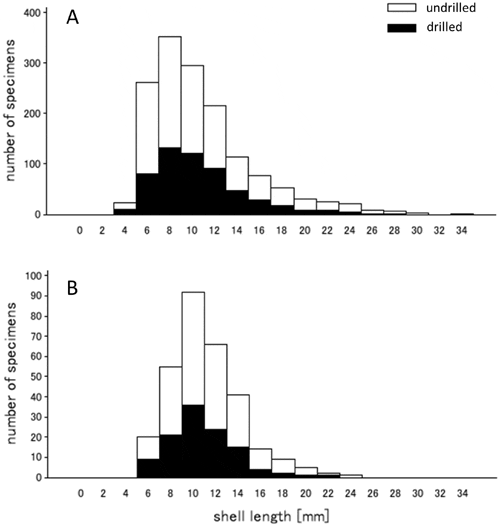
No dramatic changes in indirect ecological proxies measurable from shell assemblages could be demonstrated across seasons or through time. Whereas the absolute abundance of specimens per transect decreased notably over the last 30 years (Fig. 4), the rank abundance of the three dominant species remained unchanged regardless of the season, with C. gallina being the dominant species and D. semistriatus the least abundant species (Fig. 4). Similarly, drilling frequency, a proxy for predator-prey interactions between carnivorous gastropods and bivalves, showed consistent patterns through time. C. gallina was drilled frequently in both time intervals (48.3 per cent of drilled valves in 1978–1981 and 41.7 per cent of drilled valves in 2008–2010, respectively), D. semistriatus was drilled infrequently (8.2 per cent for 1978–1981 and 8.9 per cent for 2008–2010), and D. trunculus was drilled rarely (1.3 per cent for 1978–1981 and 4.3 per cent for 2008–2010). Seasonal changes in drilling frequency also persisted through time. For both time intervals and for each species, drilling frequencies were notably lower during summer months. Finally, comparison of size frequency distributions for specimens of C. gallina obtained for August transects did not suggest any dramatic shifts in body size between the two time intervals (Fig. 5). The monthly median values for valve length varied in a comparable range for 1978– 1980 August transects (n = 826, M1978 = 8.11, n = 205, M1979 = 12.48, n = 452, M1980 =12.30) and 2008–2010 transects (n = 259, M2008 = 10.69, n = 46, M2008 = 9.99). Moreover, the variation in median shell size was more variable between the transects within the 1978–1980 time interval than across the two time intervals. The maximum difference in medians for 1978– 1980 was 3.19 mm (M1980 vs. M1979), whereas the maximum difference in medians observed between the two time intervals was 2.58 mm (M1978 vs. M2008). When data were grouped by both, time interval and month, the two time intervals did not differ significantly from one another, once among-transect variation within time intervals was considered explicitly (Unbalanced 2-Way Anova, df =1, MS= 40.27, F = 2.85, p= 0.09).
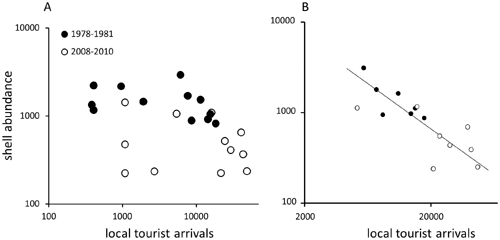
When shell abundance estimates for each transect was compared with corresponding local tourist arrivals, significant negative correlations were observed for monthly data (n= 51, r=20.52, p,0.0001) and for data averaged by month within each time interval (n= 24, r =20.49, p = 0.015). Absolute values of correlation coefficients were relatively modest and the bivariate interrelation was not strong when examined visually for data averaged by month (Fig. 6A). However, the tourist arrivals were distinctly bimodal (note a gap along x-axis of Fig. 6A). When data were restricted to months when tourist arrivals had been high (.5000), a strong negative correlation was observed (Fig. 6B), whether analyzed for monthly data (n= 32, r =20.72, p,0.0001) or averaged by month within each time interval (n= 15, r =20.84, p = 0.0001). For restricted data, tourist arrivals were a reasonable predictor of shell abundance (r2 = 0.64, Reduced Major Axis Regression, p =0.0008, Permutation Test). When data were analyzed separately for each time interval, correlation coefficients remained negative, although not always significant (as may be expected given a loss of statistical power associated with reduced sample size). Correlations between first differences (month-tomonth change in shell abundance versus month-to-month change in tourist arrivals) were also negative, but less pronounced and non-significant in most cases. Because adjacent transect estimates were separated on average by 30 days, they did not represent directly adjacent time series datapoints with high potential for strong co-dependences. Consequently, detrending may be an overly conservative and potentially misleading corrective strategy.
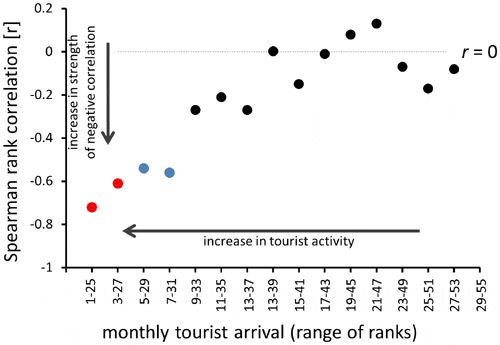
When data were grouped by tourist arrivals, the strength of correlation increased as a function of tourist arrivals (Fig. 7). The Spearman correlation coefficient reached the most negative value for data restricted to 28 months with the highest tourist arrivals and fluctuated around 0 for months with the lowest tourist arrivals (Fig. 7).
Discussion
Causes of Decline in Shell Abundance
The three models of shell decline evaluated in this study include trends in hydrodynamics resulting in changes in rates of onshore transport of shell material; ecological shifts in population dynamics and ecosystem characteristics resulting in time-variant shell input; and changes in tourism activities and tourism-related beach management practices resulting in varying rates of shell removal from shoreline areas. Whereas these models do not represent an exhaustive list of causal explanations, many other obvious factors such as changes in beach geomorphology, urban encroachment, or an increase in commercial fisheries are unlikely to apply. Other possible causal drivers are expected to be reflected in patterns evaluated by the three postulated models (e.g., an invasive species could alter rank order of dominant species, thus supporting ecological changes through time).
Seasonal changes in wave energy may result in differential delivery of shell material to the beach and decadal-scale changes may consequently produce similar changes over longer time scales. Because wave energy has remained relatively unchanged over the last 30 years and shell abundance did not show any significant correlations with wave height regardless of data groupings and tests, it is unlikely that changes in local hydrodynamic played a notable role in the observed decline in shell abundance.
Population dynamics of the three studied species may have varied seasonally or through time, resulting in a variable input of new shell material to the beach. Because the initial shell input into death assemblages is expected to be primarily controlled by biological productivity, including spawning patterns, mortality rates, and related locally-controlled processes, seasonal and multi-decadal changes in shell input at Llarga Beach cannot be evaluated directly (bivalve population data for the study area were not available). However, over the last 30 years the same three species have dominated the local shell assemblages and have shown comparable seasonal trends. It is, thus, unlikely that the observed decline in shell abundance in the last three decades was driven by changes in population dynamics. Moreover, for each of the three species size frequency distributions remained comparable through time, suggesting temporal stability in mortality and recruitment patterns. Finally, patterns of drilling attacks by predatory snails (within and across prey species) did not change notably between the two time intervals. Thus, all indirect ecological proxies measurable from the sampled shell assemblages consistently suggested that no major changes took place in local ecosystems over the last 30 years.
A significant negative correlation was found between tourism and shell abundance consistently at several levels: (1) over the last 30 years, shell abundance declined almost three-fold (2.62), a value which is remarkably close to the concurrent increase in local tourist arrivals (2.74); (2) seasonally, the substantial increase in summer tourism was congruent with concurrent decline in shell abundance; and (3) monthly, negative correlation between shells and tourist arrivals was observed, especially for months for which local tourist arrivals are high. The increase in strength of the correlation with increase in tourist arrivals is particularly compelling because it suggests that when tourist activity was high shell abundance decreased. Conversely, when local tourist arrivals were low, the tourism-associated shell loss was less important than other processes that may have contributed to variation in rates of shell accumulation and shell removal.
Active collecting versus other shell removal processes
The tourism-related processes that contribute to shell removal are certainly not limited to shell collecting, although collecting is an important activity along marine shorelines, as also demonstrated by hundreds of published shell guides aimed at leisurely and avocational shell collectors. However, other processes may be also important, including trampling, recreational clam harvesting, use of recreational vehicles, sand dredging for beach recovery, and other similar activities. In addition, beach maintenance, which can be expected to correlate with tourism intensity, often involves extensive cleaning and grooming of beach areas with heavy equipment. This is the case for Llarga Beach, where maintenance activities, using tractors with rakes, occurred daily during summer months of 2008–2010, but not during the 1978–1981 sampling interval. These maintenance activities may also explain the anomalously high decline observed for the 2008–2010 July-August months when comparing with the 1978–1981 time interval (Fig. 3B).
The relative importance of these various tourism-related processes is impossible to evaluate for Llarga Beach due to lack of relevant data. In fact, we are not aware of any published assessments addressing this issue for dead shell material (there exist numerous studies on subtidal harvesting of live mollusks for curio trading or impact of other tourism-related processes on beach and intertidal communities). The results reported here suggest that an increase in tourism can trigger a major decrease in shell abundance along shorelines thus providing justification for future studies exploring specific processes that contribute to removal of shells from beaches by tourists.
As mentioned above, other human-related activities such as commercial fisheries are unlikely to have played a significant role in the study area. Although C. gallina and D. trunculus are commercial species, fisheries along this coast have been operating over the last three decades in areas located far away from Llarga Beach (Castello´ , F., 2012, pers. comm).
Local Shell Removal as a Proxy for Global Trends
Over the last 30 years, global tourism has grown four-fold, from ~250 million tourist arrivals in 1980 to ,billion arrivals in 2010 (UNWTO Tourism Highlights 2010 by the World Tourism Organization). The nearly three-fold increase in tourist arrivals at Llarga Beach is thus not unusual. Given this increase in intensity and diversity of tourism-related processes in coastal habitats, it is likely that shell abundance has decreased on many marine shorelines, paralleling shell losses due to curio trading. In fact, some shell-rich countries that experience intense tourism have already recognized the severity of this problem and explicitly regulate not only the type, but also the quantity, of shell material that visitors are allowed to export out of the country (e.g., the Commonwealth of Bahamas). However, we lack assessments of impact of tourism on shelly organisms over multi-decadal time scales largely because of the absence of methodologically comparative samples from past decades. Estimates reported here may serve as a starting point toward developing empirical estimates for shell loss due to tourism.
It is likely that the nearly three-fold decline in shell abundance observed at Llarga beach is a conservative estimate compared to other beaches visited by tourists. While a popular tourist area, Llarga Beach is not a highest-tier tourist destination and, as noted above, the increase in tourist arrivals observed for Llarga Beach was well below global estimates for the same time interval. Llarga Beach provides shell material dominated by small, common bivalves that are neither attractive to professional shell collectors nor spectacular enough to attract attention of every casual beachcomber. Compared to other beaches, which may harbor esthetically appealing specimens, Llarga Beach is unlikely to trigger enthusiastic collecting by tourists or motivate intense subtidal harvesting of live fauna for curio trading. Finally, the negative effects of collecting live specimens for subsistence and recreational shellfish harvesting have been minimal in the study area compared to many other marine shorelines.
Eco-Environmental Consequences
Removal of shell material from shorelines may trigger negative eco-environmental changes. Among others, such changes may include physical, chemical, and biological alterations.
For example, shell material scattered along the shorelines may stabilize sediments by forming pavements that hamper sediment movement, although eolian transport of fine fraction may be enhanced locally around coarse objects such as shells. Selective removal of macroscopic shells may change the rates and patterns of sediment erosion along the shoreline.
Similarly, removal of shells potentially changes calcium carbonate budget. The carbonate skeletons of marine macroorganisms such as echinoderms and mollusks tend to be overlooked in modeling the global CaCO3 cycles in the oceans, although their role may be significant. The importance of mollusks in carbon cycle has been also acknowledged by listing them as a potentially important target for carbon sequestration. As our understanding of the role of mollusks in the global elemental cycling continues to improve, we will also need to account for the removal of large quantities of mollusk shells from the natural cycle by tourism. If the removal of shells due to tourism along shorelines increased three-fold over the last three decades, as this study potentially implies, the tourists may be significantly altering the CaCO3 cycling on our planet.
Removal or destruction of shells, which serve variety of ecosystem functions, may also negatively affect a broad spectrum of organisms. The obvious examples include use of shells for anchoring by seagrass, settling for diverse encrusting organisms, and dwelling and mining by endobiolithic algae and bioeroding sponges. Hermit crabs and fish also rely heavily on mollusk shells (mostly gastropods) and are often limited by lack of suitable shells. Shells also serve as building materials for variety of coastal organisms, both terrestrial and marine, and perform many other ecosystem services to a diverse array of organisms. It is fair to note here that the increase in shell abundance can also have negative effects. For example, beetles of the genus Bledius do not live in sand too rich in shells because of their digging behavior and sediments with a high fraction of shell fragments may negatively affect intertidal clams.
Conclusions
By exploiting a unique opportunity for developing comparative data on shell abundance for one specific coastal area on the Iberian Peninsula, the study demonstrated that shell abundance declined substantially over the last three decades. Moreover, the shell abundance patterns showed strong negative correlations (over multiple time scales) with tourist arrivals suggesting that tourism and tourism-related activities may be a driving force behind the accelerating removal of shells from marine shorelines. The shell loss may have substantial ecological and environmental consequences, although a rigorous quantitative assessment is not yet possible.
There is a growing realization that the most recent fossil record, including surficial skeletal remains found on land and in the sea, may yield diverse data for assessing ecosystem changes. This study suggests that shell accumulations found along shorelines may also provide quantifiable information about local consequences and global implications of seasonal and multi-decadal changes in tourism.
February 2014

