The volume of global finfish aquaculture production, as well as the range of species farmed, has in - creased significantly during the last 2 decades (FAO 2011). For instance, more than 15 euryhaline marine fish species are now cultured in the Mediterranean basin. The most important cultured species are gilthead sea bream Sparus aurata and European sea bass Dicentrarchus labrax. In 2010, the total production of sea bream and sea bass was approximately 140 000 and 120 000 t, respectively (FAO 2011). In order to facilitate further production increases, prevention of infectious diseases in farm stocks is important (Salama & Murray 2011). This will implicitly reduce the potential for significant economic losses through mortality, costs of treatments and carcass removal, or decreased growth rate of moribund and convalescent fish (Murray & Peeler 2005). Despite a growing fish farming industry and accompanying research efforts, the general fish health status in Mediterranean aquaculture has been considered to have deteriorated in recent years (Subasinghe 2009). The scientific and legislative efforts to ameliorate this situation are apparently not sufficient to prevent disease propagation. Also, there is a considerable lack of empirical knowledge regarding the mechanisms that trigger disease transfer, as most of the epizootiological risk assessment studies of disease transmission in aquaculture have been simulated theoretically without experimental or in situ testing under farming conditions (Peeler et al. 2006).
The rapid expansion of fish farming into new areas might involve propagation of ‘old’ pathogens from the environment to the new cultured stocks, while new environmental conditions may favour growth and expansion of pathogens already present within the new farm stock. For example, in order to produce new and diversified products, novel species are in - tro duced to existing rearing systems, imposing rearing pressure on new fish species inclined to immunosuppression. Under such conditions, bidirectional pathogen transfer is highly possible (e.g. from the new species to the environment and vice versa), since new stocks represent a susceptible focal point of pathogen proliferation. Secondly, the intensification of fish culture, through increased production per farm unit, might also increase the concentration of pathogens within cages and thus facilitate their spreading (Murray 2009), aided by fish immuno - suppression resulting from a general adaptation syndrome in intensive rearing (Ashley 2007). In consequence, pathogen concentration in the surrounding environment can increase with the increased numbers of infected fish on a farm, irrespective of their density (Murray 2009).
Disease problems may arise because the current farming techniques and the open design of Mediterranean aquaculture systems permit transmission of pathogens among pens. The primary route for introduction of pathogens to a farm is through transport of infected seedlings from hatcheries (Piper et al. 1982), food, infected equipment, staff and vessels (e.g. Kennedy & Fitch 1990, Ruiz et al. 2000, Murray & Peeler 2005), as well as through water currents (Fouz et al. 2000, Amundrud & Murray 2009, Frazer 2009, Salama & Murray 2011). Monitoring of animal health, biosecurity programmes and other disease control measures, including disease management areas or surveillance zones around farms in site selection programmes, are designed to minimize the risk of spreading disease through these pathways (DAFF 2000, Scottish Executive 2000, Rae 2002, Subasinghe et al. 2004, Bondad-Reantaso & Subasinghe 2008, Lyngstad et al. 2008, Mardones et al. 2009, Marine Scotland 2010). The European Community basic legal provision ‘Council Directive 2006/88/EC’ lays down the minimum prevention, control and eradication measures for aquatic animal diseases to be implemented in aquaculture activity by the Member States. However, the efficiency depends on farmers following the rules. Although disease control regulations for the Mediterranean finfish aquaculture industry are well established (Le Breton 1999, Rodgers & Basurco 2009), the current legislation is not consistent throughout the Mediterranean (Cardia & Lova - telli 2007). For instance, seedling import/ export regulations and quarantine measures still vary from country to country.
In addition to the ‘traditional’ pathways for disease transmission, movements of both wild fish and farm escapees also need to be considered as vectors for disease propagation (Fig. 1). This has been studied in more temperate regions of the world (e.g. Diamant et al. 2004, Naylor et al. 2005, Raynard et al. 2007, Johansen et al. 2011), but not in the Mediterranean. The aim of the present study is to discuss the po - tential risk of pathogen transmission in Mediterranean open-sea aquaculture through movements of escapees and farm-aggregated fish, and we suggest that this should be taken into account in management strategies to ensure the sustainability of aquaculture activity in Mediterranean countries.
According to the World Organization for Animal Health (OIE 2012: www.oie.int/international- standardsetting/ aquatic- manual/ access- online/), the most frequent Mediterranean pathogens belong to secondary microorganisms that are firmly ingrained at the farm site and emerge periodically, triggered by shifts in environmental conditions or anthropogenic stress.
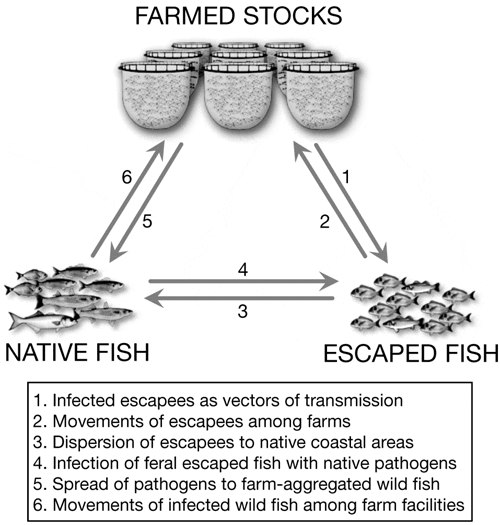
These ‘domestic’ pathogens are able to infect farmed stocks but are also present in the farm even when no clinical symptoms are observed. Such local diseases may perhaps be the most probable candidates for propagation through escaped fish as they are treated according to farmers’ experience, which in turn may increase the risk for development of resistant strains.
Escapees as Potential Vectors of Transmission
A characteristic of net-pen production units is that fish may escape from the pens (Johansen et al. 2011). Over 7 million farmed sea bream and sea bass have been reported to have escaped from Mediterranean farms in 67 small- and large-scale incidents (Jackson et al. 2012). Escapees may significantly alter the temporal or spatial distribution of pathogens occurring in farms and in the wild. Apart from the pathogen characteristics and level of fish infection within the farms, the risk of disease propagation depends on survival and post-escape behaviour of the escapees, as well as the wild fish abundance outside farms. However, the health status of escapees and the viability/pathogenicity of their putative pathogens have yet to be examined in the Mediterranean.
Several studies on escaped sea bream and sea bass in the south-western Mediterranean Sea have demon strated connectivity among farms and other marine areas of interest, like local fishing grounds (Arechavala-Lopez et al. 2011b, 2012). These findings indicate a risk of spreading diseases and pathogens through movements of escapees, both to other farms and wild fish populations. Farmed sea bream and sea bass share many pathogens with their wild conspecifics and other wild fish species, which also increases the risk of pathogen transmission (Raynard et al. 2007). Therefore, the dispersion of farm-escaped fish should be taken into account in disease control programmes.
Although there are no regulatory measures established in Mediterranean aquaculture to reduce the risk of fish escape, these do exist in several other countries (i.e. Canada, Norway, USA, Chile, Scotland). These strategies have increased our knowledge about pathogen transmission through esca - pees, in particular for salmonids (e.g. Heuch & Mo 2001, Krkošek et al. 2005, Naylor et al. 2005, Ford & Myers 2008, Thorstad et al. 2008, Revie et al. 2009, Jensen et al. 2010, Johansen et al. 2011). For instance, escaped Atlantic salmon have been identified as reservoirs of the parasitic sea lice LepeophLepeophtheirus salmonis in coastal waters (Heuch & Mo 2001, Revie et al. 2009), and escapees may spread lice to wild fish populations (Johnsen & Jensen 1994, Graham 2005, Plarre et al. 2005). Moreover, transmission of pathogens and parasites from farmed fish or escapees to wild populations may also occur in farming of Atlantic cod Gadus morhua (Øines et al. 2006, Hansen et al. 2008). In addition, connectivity among cod farms through movements of escapees has been demonstrated (Uglem et al. 2008, 2010), although direct evidence for transfer of pathogens is lacking at present. With the exception of salmon lice, knowledge about transmission of pathogens from cod and salmon farming to wild fish is limited (Johansen et al. 2011). However, the actual risk of disease transmission from cod farms is currently very low, as the cod farming industry in Norway is severely reduced due to lack of economic viability. Heuch et al. (2011) suggested that trophically transmitted parasites (larval nematodes, digenean metacercariae, cestodian plerocercoids) are the most unlikely to become a health problem for farmed cod, compared to those parasites with simple life cycles and pelagic transmission stages, such as monogeneans and trichodinids, whose direct life cycle is strongly influenced by environmental conditions (temperature, salinity, organic matter). Nonetheless, the latter parasitic groups can easily lead to marketability issues due to noticeable skin infections on farmed fish.
Cross-contagion Between Farmed and Neighbouring Wild Fish Stocks
It is difficult to evaluate the health effect of escapees on the ecosystem without taking into consideration the qualitative aspects of wild fish assemblages around farms. A range of studies have shown that farms act as fish aggregating devices (for a review, see Sanchez-Jerez et al. 2011). The influence of farms on wild fish, such as diet modification, may be detrimental for some parasite species, while these same conditions could enhance others (Fernandez- Jover et al. 2010). A wide range of pathogens can be found in a large number of farm-aggregated wild fish (Raynard et al. 2007). Cross-contagion between farmed and wild fish species with shared pathogens may occur (Diamant et al. 2004, 2007) either through movements of individual fish or through speciesspecific migrations (Horbowy & Podolska 2001, Butler 2002, Murray et al. 2003, Murray & Peeler 2005, Krkošek 2010). Connection among farms and other marine areas of interest through wild fish movements have been demonstrated both in Norway (Uglem et al. 2009) and in Mediterranean fish farms. For in - stance, 2 grey mullet species (Liza aurata and Chelon labrosus) were shown to aggregate beneath cages, feeding on waste fish food, and moving among different farming and coastal areas (Arechavala-Lopez et al. 2010). In addition, farm-aggregated bogue Boops boops were found within local fisheries landings from the nearby fishing grounds (Arechavala-Lopez et al. 2011a).
Although these wild fish species share a large number of pathogens with farmed sea bream and sea bass, knowledge regarding the potential for pathogen transfer is not unambiguous. For example, in 2 Croatian farms, the polyopisthocotylean monogenean Sparicotyle chrysophrii and the cymothoid isopod Ceratothoa oestroides have both been isolated simultaneously from farmed sea bream, sea bass and wild bogue. However, analysis of the mitochondrial DNA marker cytochrome oxidase I (mtDNA COI) demonstrated that parasites inhabiting different hosts did not belong to the same population, but cluster separately, evidencing that no transfer of 2 parasite species occurred between wild and farmed fish (Mladineo et al. 2009). Interestingly, another mono - pistho cotylean monogenean, Frurnestinia echeneis, which has only recently been found to colonize farmed sea bream (Mladineo et al. 2010), has been genetically identified (mtDNA COI and internal transcribed spacer 1, ITS1, with partial 18S and 5.8S ribosomal DNA) as a shared parasite between farmed and wild sea bream (Mladineo et al. in press). However, only a minor proportion of parasite haplotypes were shared between the 2 fish populations. Moreover, when monogenean-free fingerlings were introduced to cages, they became infected by monogenean haplotypes from the farmed bream parasite population (I. Mladineo et al. unpubl. data).
Therefore, although the distribution of fish pathogens and their pathogenicity, prevalence and incidence in wild populations must be taken into account when developing a proper disease management programme (McVicar 2004, Johansen et al. 2011), many conditions must be fulfilled before pathogen transmission takes place. It seems, however, that the transfer is more likely to occur, even for generalist parasite species, if fish are in close vicinity, as well as if the hosts (e.g. farmed and wild) are of the same species. Farming conditions (oceanography of the site, density on both sides of the net-pen) further contribute to the transfer chances, but should be carefully assessed case by case before drawing conclusions.
Potential Pathogens in Mediterranean Farmed and Wild Fish
A wide range of virus, bacteria and parasite species have been described from both wild and cultured sea bass and sea bream, some of which have negative economic impacts for the fish farming industry. Most bacteria and viruses are transmitted horizontally through water or individual contact (Raynard et al. 2007), but vertical transmission through ovarian and seminal fluids also occurs (Romalde et al. 1999). In addition, vertical transmission from infected broodstock to offspring has been indicated as a dispersal route in farmed sea bream and sea bass (Castric et al. 2001, Breuil et al. 2002).
Knowledge about transfer pathways for individual pathogens, and in particular for viruses, is very sparse. However, Panzarin et al. (2012) showed that transfer of Betanodavirus between wild and farmed fish is possible, since viruses isolated from feral and farmed fish in some cases were found to be similar. Infections caused by pathogens that are shared by both farmed and wild fish are stress-related, and derived diseases usually occur at high stocking densities (Raynard et al. 2007). Skin condition also plays an important role in the onset of some bacterial diseases, since skin lesions act as sites of pathogen entry (Raynard et al. 2007). For instance, Tenacibaculum maritimum, Vibrio algynolyticus and Mycobacterium marinum are associated with stress conditions and disruption of fish skin integrity after handling (Colorni & Diamant 1992, Balebona et al. 1998, Toranzo et al. 2005).
Monogenean parasites, inhabiting skin, gills, ex - ternal body cavities and the urogenital apparatus, might induce proliferation of secondary bacterial infection between hosts (Mladineo 2002). These parasites also include generalist species, which increase the possibility of transfer between farmed and wild fish (Wootten 1989, Sasal et al. 2004). For more specialised species, host-specificity may be lost in aquaculture (Noga 2000). For instance, Mladineo & Maršic´ -Luc? i ´c (2007) reported host switch of 2 monogenean species, Lamellodiscus elegans and Sparicotyle chrysophrii, between cage-reared sparids, sea bream and sharpsnout bream Diplodus puntazzo, indicating a broadened host range. Moreover, recent colonization of farmed sea bream by the ‘new’ monogenean species Furnestinia echeneis that suppressed the generalist L. elegans from the same host, indicate that a novel pathogen has been transferred from the wild or has been imported by bream fingerlings. Transfer from the wild is plausible, since the genetic framework of F. echeneis populations in the farmed host originated from ancient haplotypes present in the wild (I. Mladineo et al. unpubl. data).
In addition, high stocking provides optimal conditions for isopod parasites. The most common isopod parasite, Ceratothoa spp., primarily isolated from farmed sea bass and sea bream, has become a problem in Mediterranean cage aquaculture (Athanassopoulou et al. 2001, Papapanagiotou & Trilles 2001, Mladineo 2003, Bouboulis et al. 2004). It has been demonstrated that infective stages of the isopode (pulli) have a time-limited natatory potential after being released from the female (Mladineo & Vali ´c 2002, Mladineo 2003), indicating that as the distance between hosts de creases, or fish density increases, the infection rate of the parasite increases. However, none of the cymothoid species reported on farmed bass and bream are known to parasitize wild conspecifics, although they occur on wild congenerics (e.g. sparids). While wild marine fish are hosts to a wide range of parasites, sometimes the dominant parasite in culture is either rare or absent in the same species in the wild, but if the dominant parasite species in aquaculture is present in wild fish populations, adverse effects are more obvious in farmed fish (Nowak 2007).
Nevertheless, the assumption that farms amplify pathogen number has recently been shown not to be valid in tuna (Mladineo et al. 2011). Although tuna farming is capture-based and relies on juvenile and adult wild tuna in contrast to sea bream/ bass farming, the latter authors observed the disappearance of monoxenous copepods and monogenean species with direct life cycles, as well as a significant decrease of heteroxenous digeneans propagated through trophic pathways. Mladineo et al. (2011) concluded that it is difficult to postulate the combination of factors affecting these parasite pop - ulations, but environmental, anthro pogenic or host intrinsic influences have to be taken into account. This finding suggests that no general rule can be applied for every aquaculture system, fish species or inhabiting parasite community, and that strict disease management plans are important.
A large number of myxozoan species have also been shown to infect farmed sea bream and sea bass, but the exact transmission patterns are unknown in most of the reported cases. For instance, Sphaerospora testicularis, which occurs in male gonads of sea bass and can cause complete destruction of the organ (Alvarez Pellitero & Sitjà-Bobadilla 1993, Rigos et al. 1999), has been detected in wild, farmed and escaped sea bass (e.g. Sitja-Bobadilla & Alvarez Pellitero 1993, Toledo-Guedes et al. 2012) but cross-contagion has not been demonstrated. However, it is suggested that S. testicularis-infected escapees could alter the reproduction dynamics of local populations after mass escape events (Toledo-Guedes et al. 2012). The sparid myxidiosis agents Enteromuxum scophthalmi, E. leei, Myxidium fugu and Myxidium sp. are transmitted spontaneously between fish (Redondo et al. 2004), and it is possible that wild fish act as reservoirs of this disease (Raynard et al. 2007). Due to low specificity, ease of fish-to-fish transmission and high pathogenic potential, sparid myxidiosis poses a serious risk both for farmed and feral fish communities (Raynard et al. 2007).
Transmission of bacterial diseases among wild grey mullets (e.g. Mugil cephalus, Chelon labrosus, Liza spp.) and sparids (e.g. Boops boops, Sarpa salpa, Diplodus spp.) and farmed fish is well documented. For example, transmission of Streptococcus iniae and Mycobacterium marinum from sea cages to wild fish in the Mediterranean and Red Sea has been confirmed (Diamant et al. 2000, Colorni et al. 2002, Ucko et al. 2002, Kvitt & Colorni 2004). Furthermore, the same strain of S. iniae identified from infected farmed fish was isolated from wild fish as far as 2 km from the cages (Colorni et al. 2002), and the same strain of S. agalactiae was isolated from cultured sea bream and wild mullet Liza kluzingeri in the Arabian Gulf (Evans et al. 2002). Unlike parasitic pathogens, bacteria seem to have a higher potential to spread between wild and farmed fish, probably because the ecological barriers that exist for parasite transfer do not represent a great obstacle for bacteria. Firstly, bacteria are almost always present on fish surfaces. Secondly, bacterial diseases are usually treated by non-professional staff at the farms, consequently involving increased risk of developing resistance and more pathogenic strains. Finally, bacteria are often generalists and do not need wild conspecifics to spread from farmed fish.
Guidelines and Conclusions
There is a potential risk of pathogen transmission through movements of escaped and wild fish in Mediterranean fish farming areas, but actual transmission has been documented only in a handful of cases. Infected farmed fish that escape from cages by technical or operational failures may in theory spread pathogens to other cages/farms, as well to wild fish (Arechavala-Lopez et al. 2011b, 2012). In addition, infected wild fish might transfer pathogens back to the farmed fish, or become infected by farmed stocks if they stay close to farms (Raynard et al. 2007). Moreover, the large variety of shared pathogens among wild and farmed fish and the various pathways of pathogen transmission increase the potential for infection and render epidemiological risk management difficult. Therefore, disease outbreaks at farms could facilitate the transmission of pathogens from fish which are held in great numbers in cages and can easily be dispersed through water currents and aforementioned fish movements, finding a potential host. Further research on molecular mechanisms of disease transmission in aquaculture and marine environments, as well more holistic analyses of pathogenic events in the Mediterranean, are needed to clarify the potential of transmission of pathogens from aquaculture to the marine ecosystems.
As Mediterranean aquaculture is expected to in - crease in the future, actions that reduce the risk of disease emergence, establishment and spread are required. There are several factors involved in pathogen transmission that should be taken into account in this context, for example host density, pathogen-host specificity, proliferation of disease and environmental characteristics. As for other forms of animal health management, preventive measures are the most effective, cost efficient and long lasting (Meyer 1991). Firstly, improving biosecurity and disease control programmes is necessary at all levels of the fish farming industry, from the production unit to markets, in order to prevent and control diseases and preserve human, animal, and environmental health. Further, the existing spacing system for fish farms requires reconsideration based not only on knowledge of hydrodynamics and human activities but also based on information about fish movements. Unfortunately, the development of effective siting systems may be impeded by the expected increase in the farming industry and the large-scale movement patterns of many fish species along the coast.
Since the existing knowledge about pathogen transfer in Mediterranean aquaculture is sparse, there should be a greater focus on health monitoring of farmed fish as well as on prevention of escapes. Escape incidents may be reduced by improving cage technology and operational routines. Improved routines for reporting escape incidents and infectious episodes would also contribute to improved management. Management agreements among neighbouring farms are also essential tools for risk mitigation (Gustafson et al. 2007), especially if escapees are exposed to prophylactic or therapeutic treatment that could influence the health of the consumers. Finally, future risk assessments for aquaculture management should be improved, and biological interaction among farmed stocks, escapees and wild fish populations should be taken into account. In conclusion, preventive strategies, contingency plans and mitigation measures, as well as spatial planning, which take into account the risk of disease transmission to wild populations and fishing grounds, are essential for sustainable development of Mediterranean aqua - culture. Because Medi terranean aquaculture is poorly and non- uniformly regulated (Chapela-Perez & Ballesteros 2011), future management measures should be made mandatory under governmental regulations, and should be supported by local, regional or international legislation.
November 2013



