Large patches of anthropogenic marine debris have been reported from 2 of the ocean’s largest features—the North Pacific and North Atlantic subtropical gyres (Moore et al. 2001, Law et al. 2010). Plastic and other anthropogenic debris have been found in the stomachs of a large number of marine organisms, including sea birds (e.g. Spear et al. 1995), invertebrates (Graham & Thompson 2009), marine mammals (e.g. Eriksson & Burton 2003), and planktivorous marine fishes (e.g. Boerger et al. 2010). Further studies documenting plastic debris throughout benthic, pelagic, and littoral marine environments provide direct evidence that plastic pollution is widespread, persistent, and an important issue of concern (e.g. McDermid & Mc Mullen 2004, Thompson et al. 2004, Carson et al. 2011, Doyle et al. 2011).
The detrimental effects of plastic debris on marine biota include physical entanglement, decreased nutrition from intestinal blockage, and suffocation or decreased mobility (Derraik 2002, reviewed in Gregory 2009). Several studies have detailed ecotoxic effects of plastic debris in the marine environment, whereby plastic can serve as a vector for harmful organic contaminants such as polychlorinated bi phenyls, aromatic and petroleum hydrocarbons, and organochlorine pesticides (reviewed in Teuten et al. 2009).
Despite the prevalence of studies documenting the environmental implications of plastic debris in the world’s oceans there have been few reports of plastic ingestion by large marine fishes. Two recent studies from the North Pacific Subtropical Gyre reported microplastic in the stomachs of planktivorous micronekton fishes, with approximately 9 to 32 per cent of fishes examined containing plastic (Boerger et al. 2010, Davison & Asch 2011). Reports of plastic ingestion in larger, higher trophic level pelagic fishes are sparse in the literature but include isolated reports from species such as dolphinfish, tunas, and the southern ocean moonfish (Hoss & Settle 1990 and references therein, Jackson et al. 2000). We investigated ingestion of plastic and other anthropogenic debris in 7 species of large pelagic predatory fishes from open ocean waters of the central North Pacific Subtropical Gyre surrounding the Hawaiian Island Archipelago. Our study was based on incidental obser vations collected from multiyear diet studies, the primary ob - jective of which was to describe regional food habits and trophic ecology. Our findings highlight 2 novel perspectives of the pervasive global plastic debris issue, namely the presence of plastic in some commonly consumed pelagic fish species and the suggestion that plastic pollution continues to extend into the deep ocean through interconnected epi- and mesopelagic food webs.
Materials and methods
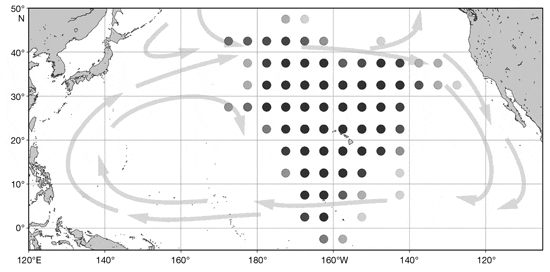
Pelagic fish stomachs were collected at sea by trained longline fishery observers of the National Oceanic and Atmospheric Administration’s (NOAA) Hawaii Observer Program (as described in Choy et al. 2009) during 2007 to 2012. Longline fishery ob servers recorded species, fish length, sex (if known), and capture date while aboard commercial fishing vessels operating in the central North Pa - cific Ocean. Fish specimens were collected from both shallow and deep longline sets, which are typically set at ~0−100 and ~100−400 m, respectively (Bigelow et al. 2006). In accordance with NOAA’s Fisheries Operations Data Confidentiality Policy, catch locations are not reported here. However, Fig. 1 details the general spatial coverage of the fishery, approximating the area from which fish in this study were sampled. Fishery observers sampled fish specimens randomly and opportunistically, and all stomach samples provided were examined in the laboratory.
Recent morphological and genetic studies suggest that opah species in the North Pacific are a complex of 2 species, referred to hereafter as Lampris sp. (bigeye) and Lampris sp. (small-eye) (J. Hyde pers. comm.). The tropical species of opah is currently known as Lampris guttatus. Stomachs of Lampris spp. are not commonly excised at sea, thus stomachs of these species were primarily sampled directly from local seafood wholesalers. While fish length and/or whole mass were available for these specimens, capture date and location were not. The large majority of Lampris spp., Gempylus serpens, Alepisaurus ferox, and Lepidocybium flavobrunneum specimens were provided whole. No evidence of regurgitation of stomach contents (e.g. food items in esophagus or gills, everted stomachs) was noted.
In the laboratory, stomachs were defrosted and contents were sorted to the lowest taxonomic level possible using published literature and the advice of taxonomic experts. Ingested items (both prey and anthropogenic debris) were weighed to the nearest tenth of a gram, and their lengths measured if intact enough to do so. Stomach content examination methods were capable of detecting all visible size spectra of non-biological debris pieces, including microplastics (
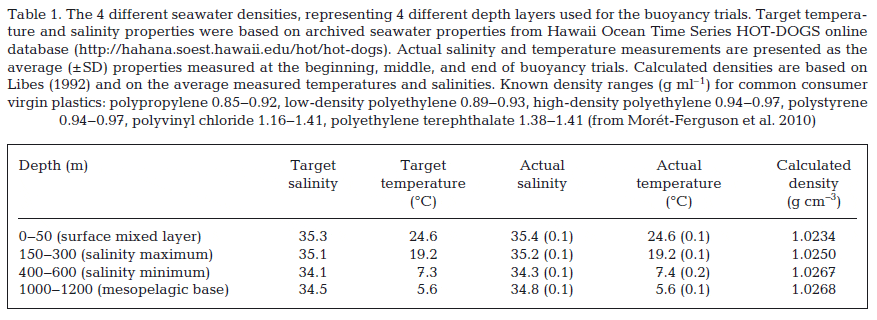
The fish species examined forage and move across different depth strata in the water column and not all are thought to come to the surface (see depth utilization data from fish tagging studies from the central North Pacific: Polovina et al. 2007, Howell et al. 2010, Abecassis et al. 2012). To investigate where in the water column debris pieces may have been ingested, the buoyancy of a representative subset of the plastic and other debris was evaluated (159 pieces from 58 individuals belonging to 3 fish species). These plastic and other anthropogenic debris pieces were weighed to the nearest hundredth of a gram, measured in the longest and widest dimensions to the nearest millimeter with calipers, and categorized by color and general debris type. Seawater representing 4 densities was made up using Instant Ocean sea salt and a controlled-temperature water circulator. Temperature and salinity properties representative of 4 different depth layers in the North Pacific Subtropical Gyre were extracted from the Hawaii Ocean Time Series HOT-DOGS online database (Table 1). 1.5 l of each seawater type was placed into 2 l glass beakers within controlled-temperature water baths; water was replaced halfway through each buoyancy trial (i.e. after approx. half of the subset of debris pieces were floated in that water type) to prevent potential changes in physical properties. Debris pieces were inspected for visible air bubbles and if present air bubbles were removed. Each debris piece was floated in each of the 4 water types for 60 s. Pieces were dipped in distilled water to avoid crosscontamination between water types. Every 10th piece of debris was floated twice in each of the 4 water types; no buoyancy changes were observed across replicates. Temperature and salinity were measured at the start, middle and end of each trial for the 4 water types using a handheld conductivity meter (Orion 135) (Table 1).
Results
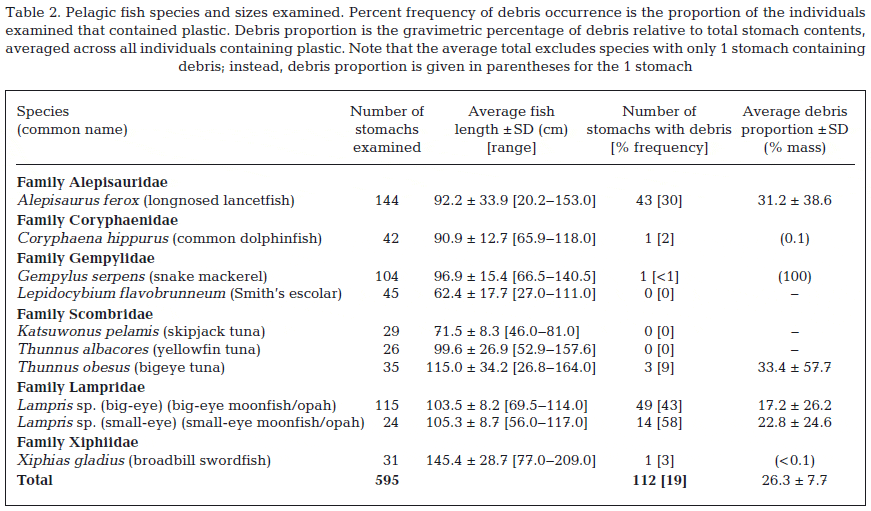
The stomach contents of 595 individuals encompassing 10 species (6 families) were identified. Debris of anthropogenic origin was found within the stomachs of 112 individuals, i.e. 19 per cent of all fishes examined (Table 2). Percent frequency of ingestion among all 10 species ranged from 0 to 58 per cent, and was highest in the 2 Lampris spp. (43 per cent of Lampris sp. (big-eye) and 58 per cent of Lampris sp. (small-eye)) and Alepisaurus ferox (30 per cent), all presumably mesopelagic fishes (Polovina et al. 2007). Of the 10 fish species examined, 3 contained no debris of anthropogenic origin.
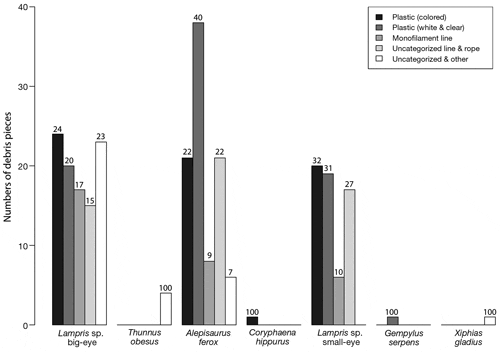
Of the 7 species of fishes that in - gested marine debris, anthropogenic items consisted mainly of assorted pieces of plastic and different types of fishing and nautical lines and rope pieces. A total of 262 individual pieces of plastic and other marine debris were found, and examples of some of these different types of debris are shown in Fig. 2. Categorical debris composition by species is shown in Fig. 3. Of the 3 species that ingested the largest number of debris pieces, Alepisaurus ferox seemed to favor white and clear plastic pieces while the 2 Lampris species did not seem to favor one type of debris over another (Fig. 3).

Within the debris subset of 159 individual pieces, debris length, width, and mass ranged widely (Table 3). On average Lampris sp. (small-eye) had one of the highest incidences of debris ingestion, and of the 3 species examined in ferox, 45 per cent for Lampris sp. (big-eye), and 31 per cent for Lampris sp. (small-eye)). The second most common color of ingested debris was black (14 per cent for A. ferox, 22 per cent for Lampris sp. (big-eye), and 31 per cent for Lampris sp. (small-eye)).
All 159 debris pieces available for buoyancy trials floated in all 4 types of sea water representing depths of 0−50, 150−300, 400−600, and 1000−1200 m (Table 1). All pieces were floated as described above for 60 s per 4 water types, however we note that 2 of each type of debris (categories as described in Fig. 3) were left in each water type for longer than 10 min and all pieces remained positively buoyant. Furthermore, different types of debris were physically forced to the bottom of the beaker for all 4 water types and each piece immediately returned to the surface of the water.
Discussion
Very few previous studies have specifically investigated debris ingestion in large marine fishes, but rather have documented such ingestion incidental to conventional food habit studies. Previous studies reporting plastic debris ingestion have focused on juvenile, coastal, and mesopelagic micronekton marine fishes (e.g. Carpenter et al. 1972, Boerger et al. 2010, Davison & Asch 2011), all very distinct from the large, higher trophic level pelagic fishes studied here. A handful of previous studies have reported on plastic debris ingestion in trophically similar predatory meso pelagic fishes, however observations from most of these studies are based on relatively small sample sizes. For example, Jackson et al. (2000) examined 69 Lampris immaculatus individuals from the Patagonian Shelf and found plastic in 14 per cent of the fishes. Kubota & Uyeno (1970) reported 78 individual plastic pieces in the stomachs of 36 Alepisaurus ferox specimens, and Fujieda et al. (2008) reported plastic in 1 A. ferox individual. Our study reports observations of frequent plastic ingestion based on almost 600 individual fishes encompassing 10 species. With these considerable sample sizes we report some of the highest incidences of debris ingestion for similar marine fishes, with 30 per cent of A. ferox, 43 per cent of Lampris sp. (big-eye), and 58 per cent of Lampris sp. (small-eye) individuals having ingested some form of anthropogenic debris (Table 2). These observations suggest that debris ingestion is not uncommon and may be more prevalent in certain species than previous observations have suggested.
The 3 species with the largest sample sizes and highest ingestion frequencies (Alepisaurus ferox and Lampris spp.) are considered to be primarily mesopelagic based on limited tagging and diet infor - mation. Polovina et al. (2007) described the diving behavior of 11 Lampris spp. (formerly L. guttatus) around Hawaii as inhabiting ~50−150 m at night and ~100−400 m during the day, with the shallowest and deepest depths recorded as 28 and 736 m respectively. There are no known tagging studies of A. ferox in the central North Pacific Subtropical Gyre, but the most important prey items from diet studies are primarily mesopelagic micronekton, including many non-migratory species (e.g. paralepidid and sternoptychid fishes, hyperiid amphipods) (Haedrich & Nielsen 1966, Choy 2008). Thus, a central question arising from our findings is why mesopelagic predatory fishes are feeding on plastic debris of a presumably surface ocean origin.
There are several possible explanations for finding plastic pieces in the stomachs of fishes that forage primarily in the mesopelagic zone. First, the tagging and diet studies have incompletely characterized the movements of these fish and they do in fact at times feed in surface waters at night. The second is that the debris pieces are found in the predator’s guts because their prey had first consumed them (i.e. secondary ingestion, as discussed in fur seals by Eriksson & Burton (2003)). We can quickly discount this possibility because the plastic particles consumed by micronektonic fishes (i.e. the prey of Lampris spp. and Alepisaurus ferox) are typically ~4 cm, Table 3) (Boerger et al. 2010, Davison & Asch 2011). In fact, many of the plastic pieces we found were as large or larger than other prey found in the guts of the predators (Choy 2008).
Another explanation is that the plastic and other debris were actually consumed at depth. As expected for plastic debris found offshore of landmasses, all plastic and other debris pieces were positively buoyant. There was no visible evidence of encrusting fauna that would have affected the inherent buoyancy of individual debris pieces. However, buoyancy was evaluated following consumption by the fishes and it is possible that buoyancy properties changed after ingestion. Buoyant plastics are known to sink when waterlogged and/or biofouled (Ye & Andrady 1991, Barnes et al. 2009, Morét-Ferguson et al. 2010, Lobelle & Cunliffe 2011). Plastics are found on the seabeds of all major oceans including the abyssal plains (Galgani et al. 1996, 2000, Barnes et al. 2009, Ramirez-Llodra et al. 2011, Bergmann & Klages 2012) and are the most common debris found along the slope of the eastern Pacific (Keller et al. 2010). Thus it is reasonable to assume that plastics are present in the water column either as neutrally drifting pieces or slowly sinking particles. However, biofouling organisms would be digested in an acidic fish stomach after ingestion and the debris buoyancy properties would change. Davison & Asch (2011) found that non-migrating planktivorous fishes ingested plastic, providing further suggestion that there is plastic in deep midwaters.
Finally, it is possible that wind-driven mixing, down welling and/or currents act to carry plastic particles below surface waters, enabling ingestion by mesopelagic fishes. Kukulka et al. (2012) showed from trawl samples and mixing models that buoyant millimeter-sized particles of plastic were distributed throughout the upper wind-mixed water layer. Whether the larger particles identified in this study could also be physically mixed to depth remains uncertain.
Why fish would consume plastic or other types of marine debris is uncertain. It is often hypothesized that seabirds and marine turtles confuse plastic particles with gelatinous prey (Carr 1987, Gramentz 1988) and the small plastic particles in zooplanktivorous fishes were predominantly blue, clear, and white, and of similar size to their main prey (Boerger et al. 2010). In our case, the differences in prevalence of plastic across species may afford some clues. Al - though they had the lowest sample sizes, the species that seemed to have the lowest incidences of plastic ingestion include Thunnus spp., Xiphias gladius, and Coryphaena hippurus. C. hippurus is a neustonic predator feeding predominantly on neustonic fishes, some squids and crustaceans (Olson & Galván-Magaña 2002). Thunnus spp. and X. gladius principally consume active micronektonic squids and fishes (Holland & Grubbs 2007, Choy 2008, Watanabe et al. 2009). While Lampris spp. also consume many micronekton fishes and squids (e.g. Jackson et al. 2000), specimens from the central North Pacific are known to also frequently prey on hyperiid amphipods (C. A. Choy et al. unpubl.). Alepisaurus ferox is also known to consume large numbers of mesopelagic hyperiid amphipods (Satoh 2004, Potier et al. 2007), namely the species Phrosina semilunata, Phronima sedentaria, and Platyscelus spp. identified from a study from the North Pacific (C. A. Choy et al. unpubl.). Many hyperiid amphipods associate with or are parasites of gelatinous zooplankton (Brusca 1973, Harbison et al. 1977, Gasca & Haddock 2004, Gasca et al. 2007). As gelatinous prey are difficult to identify in stomach contents due to rapid digestion rates, it is unclear as to whether predatory fish consume just the amphipods and/or their gelatinous hosts. There is evidence that A. ferox commonly ingests gelatinous hosts (Satoh 2004, C. A. Choy et al. unpubl.) and thus it is possible that the 2 Lampris species and A. ferox confuse the plastic pieces for gelatinous prey. Additionally, almost half of the debris pieces found in the stomachs of A. ferox and the 2 Lampris species were transparent, translucent or white, providing a visual basis for possible confusion with common pelagic gelatinous animals such as salps or siphonophores.
While we focus on the differences in the prevalence of plastic and marine debris in shallow versus deeper-dwelling pelagic fishes there is certainly evidence for ingestion of marine debris by epipelagic predatory fishes (e.g. Coryphaena hippurus (Manooch et al. 1984), Thunnus albacares and T. atlanticus (Manooch & Mason 1983)). Thus it is possible that the apparent positive buoyancy of the debris in this study indicates surface ingestion by mesopelagic fishes, perhaps during movement into shallower waters at night as suggested from very limited tagging data (Polovina et al. 2007, Kerstetter et al. 2008).
However, considering the relatively rare incidences of debris ingestion in epipelagic versus mesopelagic predatory fishes in this study and others, it is likely that debris pieces were ingested at subsurface depths. For example, Manooch et al. (1984) examined over 2000 epipelagic C. hippurus specimens and reported percent frequencies for anthropogenic debris below 1 per cent.
Our findings highlight 2 novel perspectives of the global plastic debris issue, namely the presence of plastic in some commonly consumed pelagic fish species and the suggestion that plastic pollution continues to extend into the deep ocean through interconnected epi- and mesopelagic food webs. Furthermore, we show that plastic ingestion in large pelagic fishes is more prevalent than previously suggested. The effects of plastic ingestion on the health of the fish remain uncertain, but could include gut blockage, particularly when large pieces are consumed (as in Fig. 2) and the transfer of chemical toxins (Teuten et al. 2009). Many plastics adsorb PCBs, organochlorine pesticides, polycyclic aromatic hydrocarbons, metals, and petroleum hydrocarbons, some of which may desorb in acidic stomachs resulting in uptake to the animal (Teuten et al. 2009, Van et al. 2012). Indeed, it has been shown that seabirds that ingested plastic had higher PCB concentrations in their fat tissues (Ryan et al. 1988), and seabird chicks fed plastics showed increasing PCB concentrations (Teuten et al. 2009). Given the global commercial importance of Lampris spp. and other large pelagic fishes (namely tunas and billfishes), future research might evaluate whether these fishes carry elevated chemical toxin burdens that may ultimately pose a risk to the seafood-consuming public.
January 2014

