Anthropogenic climate change is already affecting biological and ecological systems worldwide and several kinds of responses such as phenological, biogeographical and abrupt ecosystem shifts have been documented. Kelp forests, highly productive and emblematic ecosystems of both cold-temperate and polar coastal regions, may be substantially affected by global warming. The species Laminaria digitata is among the most commercially important species of European kelps and some populations have declined for the past few years. Since the 19th century, they have been exploited by the iodine industry and more recently they have been harvested for their content in alginate, which is used as a binder, emulsifier and as a molding material in a broad range of products.
The distribution of L. digitata on the European coasts ranges from the southern coasts of Brittany (Quiberon bay) to the Northern part of the Norway. The macroalga is absent from the Baltic Sea and is sparsely distributed on the southeastern coasts of Brittany. It is found in the upper sub-littoral fringe in sheltered or moderately exposed sites and exclusively on rocky shore. It supports high hydrodynamics and prefers clear or slightly turbid waters. The temperature is a primordial factor regulating the horizontal extension of L. digitata. Its thermal optimum ranges from 10 to 15°C, reproduction is impaired beyond 18°C and death occurs when temperature reaches 22°C. Bolton and Lu¨ning found a value between 22°C and 23°C (with cell damages at 23°C). The vertical extent of this species varies according to light penetration and is therefore inversely related to turbidity. In general, the macroalga develops between +1 and 23 m but can be exceptionally found down to 20 m in very clear waters. The main competitor of L. digitata is the species Sacchoriza polyschides. This annual species grows more rapidly than L. digitata and usually colonize spaces previously occupied by L. digitata.
The decline in the abundance of L. digitata along the European coasts have gained attention recently. This phenomenon has been observed in Brittany, in Normandy and in the English Channel. Although exploitation may have contributed to the decline of the species, the implication of global warming is possible. To examine the potential influence of global climate change on L. digitata, we modeled the ecological niche (sensu Hutchinson) of the macroalga and projected its potential distribution along the European coasts for the 21st century. Such large-scale projections of changes in this marine forest canopy have never been addressed despite the fact that degradation of these habitats may have considerable ecological and economic consequences. To our knowledge, only three previous studies have applied Ecological Niche Models (ENM) on kelps: a predictive model of subtidal kelp forest in Brittany-France and two spatial predictive distribution models of the kelp species Laminaria hyperborea and Saccharina latissima in Norway. All of them used Generalized Additive Models on a small area and did not investigate the effects of climate change on the whole species distribution.
In the present study, we apply a new ENM based on presenceonly data and called Non-Parametric Probabilistic Ecological Niche model (NPPEN). This model offers many advantages: (i) it requires presence-only data, (ii) it is based on a nonparametric test and is therefore distribution-free, (iii) by the use of the generalized Mahalanobis distance, the model takes into account the correlation between environmental variables. As for any ENM, this method has assumptions and limitations, which will be later discussed.
This model, recently applied to marine fish and the macro-benthic fauna, is used to identify the ecological niche (sensu Hutchinson) of L. digitata and to model its current spatial distribution, taken as reference the period 1982–2009. We then evaluate the possible implications of global climate change for the geographical patterns of the species in the 21st century using climatic scenarios from the Coupled Model Intercomparison Project phase 5 (CMIP5). These scenarios, called ‘Representative Concentration Pathways’ (RCP), are the latest generation of earth system models designed in the framework of the Fifth Assessment Report by the Intergovernmental Panel on Climate Change (IPCC AR5). Four greenhouse gas concentration trajectories were chosen by the IPCC: RCP2.6, RCP4.5, RCP6.0 and RCP8.5 (from the most optimistic to the most pessimistic). They are labeled according to their total radiative forcing in 2100.
Results and Discussion
Selection of the Best Combination of Environmental Variables
Table 1 summarizes the analysis for each of the twelve combinations of abiotic variables tested and shows that the run based on bathymetry, maximum annual SST and salinity (experiment 2.3) presents the highest AUC value (AUC = 0.82±0.02). Therefore, we used this triplet of environmental factors to evaluate the present and the future distribution of L. digitata. However, it is important to note that the number of factors regulating the observed distribution in nature is much larger. Although here we took into account three parameters, we caution that on local basis, some parameters may be important. Interspecific interaction, the strength of local wind and its effects on wave and currents, sediments type, are some examples of parameters that may locally improve our projections.
Modeling of the Ecological Niche of L. digitata
The niche was modeled by applying the model NPPEN on the set of explanatory variables listed below: bathymetry, maximum annual SST and salinity. Probabilities of occurrence higher than p = 0.05 were found at bathymetries between 0 and 12 m (mode between 0 and 5 m), at maximum annual SSTs ranging from 1015°C to 2015°C (mode around 15°C) and at salinities between 32 and 36 (mode around 34–35), a result in agreement with the ecological knowledge of the species (Fig. S2).
Spatial Distribution of L. digitata for the Period 1982– 2009
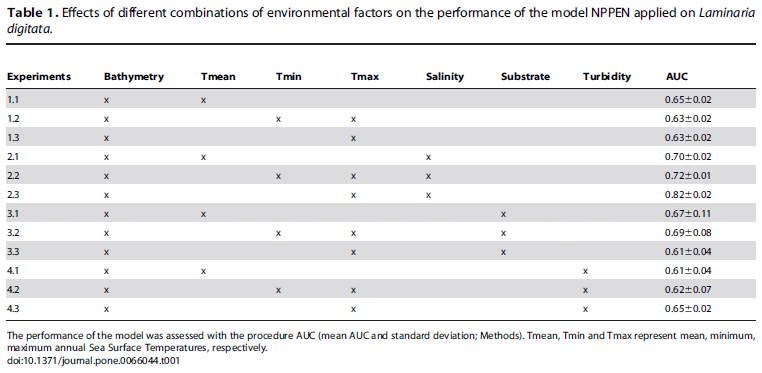
The modeled ecological niche was subsequently projected onto the geographical space to map the current distribution (1982– 2009) of L. digitata in term of probability of occurrence (Fig. 1a). A high probability of occurrence corresponds to an environment highly suitable for the species. All the southern and northern limits, the sparse distribution along the northern coasts of France, Belgium, Netherlands and Germany and the total absence of the species in the Baltic Sea were well reproduced.
Adequacy between AVHRR Data and RCP Scenarios
Since the present distribution of probability of occurrence was based on AVHRR SST data (1982–2009) and projections of future probabilities on SST data coming from RCP scenarios (2006–2100), we compared both observed and modeled SSTs for the common period 2006–2009 to prevent any bias due to the different SST datasets. This comparison was made by the use of a Taylor’s diagram. All correlation coefficients with AVHRR data were larger than 0.9, which indicate that RCP SSTs were close to AVHRR data for the overlapping period 2006–2009. These analyses suggest that any significant changes between observedand projected-SST-based probabilities of occurrence are unlikely to be related to the difference between SST datasets (Fig. S3).
Projections of the Spatial changes in the Distribution of L. digitata for the 21st Century
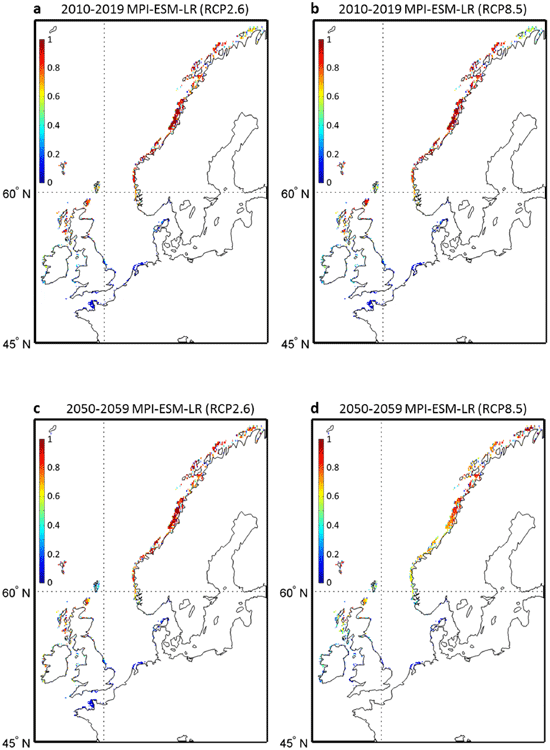
Applying a multi-model and multi-scenarios approach, we used SST data originating from the new IPCC Representative Concentration Pathways (RCP) scenarios to evaluate the effect of climate-induced changes in temperatures on both the spatial range and local densities of L. digitata. Using two climate models and three RCP scenarios (6 runs), we established projections of the spatial distribution of the kelp for each decade from 2010 to 2099. We first calculated the average probability of occurrence resulting from all runs for each decade to evaluate the potential changes in the both expected abundance and spatial distribution of the kelp until the end of the century (Fig. 1b–d). At the southern edge of the spatial distribution of the species, a substantial decline in the probability of occurrence is expected as early as the 2010s from French to Danish coasts and first nil average probabilities are observed in the 2050s (Fig. 1c). Although a diminution is clearly expected over Scotland, some populations (e.g. northern part of Scotland) are likely to persist until the end of this century (Fig. 1d). Extensive changes are also expected to take place in the southern coastal regions of Norway. A major poleward biogeographic movement of the core region (i.e. region with the highest probabilities) is likely to be observed along the Norwegian coasts in the next decades.
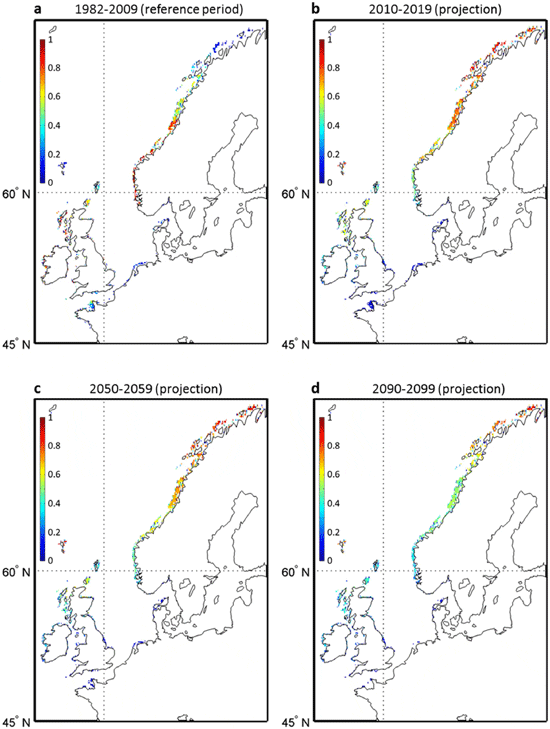
We examined the latitudinal shifts expected under three levels of warming, taken as a reference the period 1982–2009 (Fig. 2). Note that for the overlapping period 2006–2009, the probabilities calculated from observed (AVHRR) and modeled (RCP SST) data were in general close at latitudes inferior to 62uN (mean probability difference = 0.02). More differences were observed north of 62uN (mean probability difference = 0.15). It is well known that SST derived from satellite radiometric data are better evaluated in temperate areas than under high-latitudes where cloudiness related problems are recurrent. Whatever the intensity of warming, a substantial decline in the probability takes place between 45 and 62uN, as early as in the 2010s. From the decade 2020–2029 to the end of the 21st century, projected changes are subsequently smaller. With the low scenario (RCP2.6), changes taking place after 2019 are in general inferior to 0.2 (Fig. 2a) but the alterations of the latitudinal probabilities become larger when the intensity of global warming increases (Fig. 2b–c). North of 67uN, future probabilities are higher than the present ones for all scenarios.
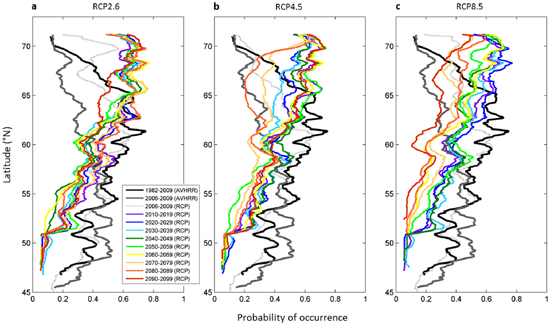
To explore the effects of both uncertainties from the two climate models and the intensity of warming from the three RCP scenarios on the range of occurrence probabilities, we calculated the per centage of runs projecting a disappearance of L. digitata during the decades 2010–2019, 2050–2059 and 2090–2099 (Fig. 3). The term ‘disappearance’ is used in geographical cells for which the probability was higher than 0.05 in the reference period 1982– 2009 and lower than this threshold for a given decade. For the 2010s, a few projections suggest some local extirpation of L. digitata from the French to the Danish coasts and in some areas from the southern parts of England (Fig. 3a) but the per centages remain low and the majority of projections only predict a regression of the populations (Fig. 1b and Fig. 2). However, as early as the decade 2050–2059, between 50 and 100 per cent of the models suggest local extirpations of this species in the previously described regions. At the end of the 21st century (decade 2090–2099), the rate of models predicting a disappearance substantially increases for most regions representing the southern part of the spatial distribution of the species and the areas in which the species is likely to become extinct spread northwards: a few projections (less than 30 per cent) even suggest local extirpation in the southern coastal regions along Norway (Fig. 3c).
Long-term Changes in the Probability of Occurrence of Laminaria digitata in Roscoff
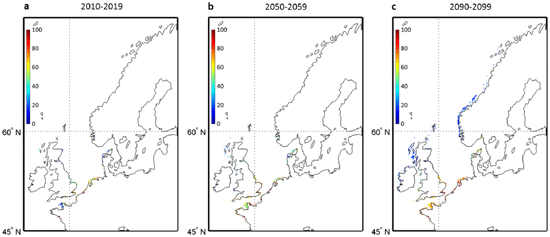
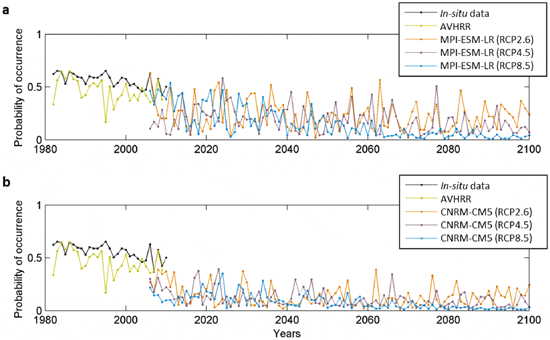
Because our projections suggest the likely disappearance of L. digitata in some countries (e.g. France) as early as the 2050s, we used local in-situ temperature data to calculate the long-term changes in the probability of occurrence of this species in an area where the seaweed is near to the southern edge of its spatial distribution (Roscoff, France) and is being exploited for its alginate and iodine content. Probabilities based on in-situ measurements were compared to AVHRR temperature-based and projected probabilities for the overlapping period 2006–2009 (Fig. 4). This analysis showed that MPI-ESM-LR-based probabilities of occurrence were closer to the in-situ-based than the probabilities based on CNRM-CM5 model, especially for scenarios RCP4.5 and RCP8.5. A long-term decline in the probability of occurrence is expected to take place in the first half of this century, mainly for scenarios RCP4.5 and RCP8.5. For Scenario RCP2.6, the intradecadal variability in the probabilities is elevated, especially for the MPI-ESM-LR model. As the CNRM-CM5 model underestimated the probabilities of occurrence based on in-situ measurements, we checked if our conclusions hold by considering exclusively the MPI-ESM-LR model (Fig. 5). We excluded from the comparison scenario RCP4.5, which gave lower probabilities. Probabilities of occurrence in the decade 2010–2019 were close to the average probabilities based on the 6 runs (see Fig. 1b). The projection based on the MPI-ESM-LR-RCP8.5 scenario suggests that extirpation of the species along French coast is very likely if global warming is extensive.
Potential Shortcomings Related to the use of Ecological Niche Models
The ecological niche is a common concept used in ecology but unfortunately, in the history of ecology, it had several definitions. In 1917, Grinnell used this term to designate the ‘‘position occupied by a species in both its biological and physical environment’’. In 1927, Elton defined the niche as the functional role of a species within the food chain. Odum, in 1953, introduced the notion of the function of organisms within a community instead of the place. Hutchinson, in 1957, defined the ecological niche as the sum of all the environmental factors acting on the organism. The niche is here a n-dimensional hypervolume and represents ‘the range of tolerance of species when several factors are taken simultaneously’. Here, as in most ENMs, we used the Hutchinson’s definition of the niche.
Ecological Niche Models such as NPPEN are although increasingly used to investigate the potential effect of global climate change on species distributions. These models have already shown their ability to produce accurate prediction of species distributions based on environmental factors. However, it is important to recall that ENMs do not take into account biotic interactions (e.g. predation, competition and mutualism), dispersal mechanisms and life-history strategies of species. Moreover, these models assume no change in the fundamental niche of species. However, rapid changes in physiological rate (especially thermal tolerance) of L. digitata can be excluded since the time-scale of this process is largely greater than a century.
Our study provides evidence that the habitat of L. digitata along the European coasts, which had suitable environmental conditions for the macroalga in the recent past have been altered by an increase in temperatures. Regional extinction in L. digitata may occur as early as the first half of this century, although this figure depends upon the magnitude of future warming and its interaction with regional climatic variability. Although local extirpation may arise directly through physiological stress related to elevated temperature, it is as likely that the retreat occurs through an unbalance in the biotic interaction between the kelp and other species, and it is plausible that the species disappears through mass mortality related to disease. In addition to global climate change, L. digitata is being affected by multiple stressors related to human activities: increased turbidity, sea level rise, introduction of exotic species. Harvesting of this species will potentially lead to the fragmentation of their populations and it is probable that the interactive adverse effects of climate change with more direct anthropogenic stressors accelerate the retreat of the kelp by reducing gene flows and genetic diversity. Several studies showed that the removal of L. digitata has often been accompanied by a concomitant increase in Saccorhiza polyschides, a pioneer noncommercial kelp species, which may have important adverse consequences not only for the associated biodiversity but for goods and services. Our longer-term projections are rather pessimistic for the future of the kelp in the English Channel and the North Sea. Any reduction of the spatial distribution of L. digitata will have important ecological consequences since the macroalga enables the establishment of a large number of animals, typically between 32 and 107 and provides spawning, nursery and feeding grounds for many species of both invertebrates and fishes.
January 2014
