Marine reserves (MR), also known as no-take marine protected areas (MPA), are widely acknowledged as a conservation tool and their utility in a variety of situations is well established . In particular over-exploited fish populations are shown to recover in the absence of fishing and generally become more abundant and attain a larger mean size in the reserve. MPAs are also frequently promoted for the management of fisheries, even though compelling evidence in support of a net fisheries benefit is lacking. Fisheries are proposed to benefit from reserves through increased production of eggs and larvae from the reserve (recruitment effect) and the net movement of adults into adjacent fishing grounds (spillover effect).
In this study we focus on the spillover effect and, to avoid confusion over the use of terms, we define spillover as the net movement of fish across the boundary of a reserve into the fished ground, which would be expected to occur on the basis of fundamental physical principles of random movement. This is in contrast to net spillover benefit which involves spillover of sufficient magnitude to compensate for lost productivity due to the closure of fishing grounds, resulting in an overall benefit to the fishery through higher catch or economic yield.
Our review of the extensive literature reporting fisheries benefits reveals that there are surprisingly few empirical studies that attempt to quantify either the recruitment effect or a net spillover benefit. For example, Goni et al. claims to be the first study to demonstrate a net spillover benefit in a fishery. Harrison et al. make a similar claim with respect to the recruitment benefit of reserves in terms of larval export. Whilst spillover has been shown in several other studies, most do not accommodate the reduction in catch that results from reducing the area of the fishery, and consequently do not demonstrate a net spillover benefit.
Fishers are generally opposed to the introduction of reserves because they reduce the size of their fishing grounds, which is inferred to result in a loss of yield. Spillover is a common counter argument from reserve proponents, including Government agencies in the US, Europe and Australia, claiming that it will compensate for the lost fishing grounds to the extent that a net improvement in fisheries yield occurs.
The impact of the introduction of reserves on yield has been addressed in a number of theoretical studies (e.g.,), several of which progressively conclude that under broad assumptions well-managed fisheries should not benefit from the introduction of reserves. Hart quantifies this result to some degree by using an age-structured model, concluding that a benefit from spillover should not be anticipated unless open area fishing mortality considerably exceeds that which produces MSY.
The assumptions underlying these studies primarily concern the homogeneity of fish stocks and are reasonable for a large range of species. The obvious exception occurs in fish stocks with strong variability in spatial structure, for example where source-sink relationships exist or where reserves may result in the closure of disproportionately productive areas. Such spatial heterogeneity is the basis of traditional spatial management of fisheries, and is a well-established and understood technique.
Targeted spatial closures can be expected to benefit fisheries for selected species if the closed area is of disproportionate significance to the productivity of the species in question. Not surprisingly some models have shown that, at least under certain conditions, higher sustainable yields can be achieved with a marine reserve than without, e.g,. But despite the common demonstration that special circumstances are required to achieve a spillover benefit from reserves, the implication of these findings have received limited attention and appear to have contributed little to the international public debate over fisheries benefits and to current management policy.
In this paper we use a widely applied fisheries population dynamics model which minimises assumptions in order for the outputs to be applicable to a broad range of fisheries in non-structured environments (‘normal’ or ‘average’ fisheries). We modify this model to incorporate a MR and consider the management circumstances under which a non-specific reserve is likely to provide a benefit to the fishery. Our work highlights the effect that the degree of mismanagement under conventional fisheries management practices has on the ability of a reserve to provide a net fisheries benefit. It also investigates how this relationship changes with the rate that fish move between the reserve and the main population.
Results
Figure 1 shows an example where a 10% reserve is introduced with 5% movement out of the reserve (μ) and a maximum growth rate (r) of 10%. This figure explores the effect of a reserve for different levels of initial effort applied to the fishery. The maximum sustainable yield (MSY) is obtained with an effort of 0.05 (ΕMSY).
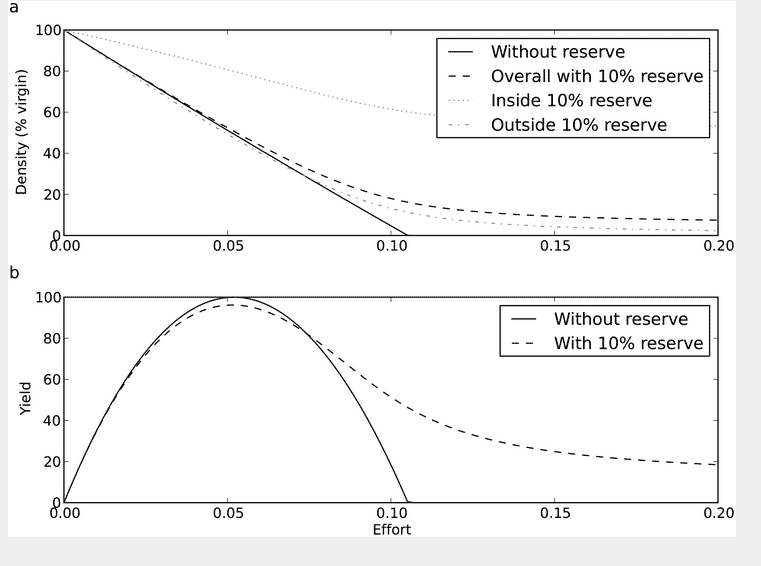
(a) The equilibrium biomass density as a function of fishing effort. The density is shown for the whole stock without a reserve and with a 10% reserve. For the reserve scenario the density inside and outside of the reserve is also shown. (b) Yield as a function of fishing effort both with and without a reserve.
doi:10.1371/journal.pone.0107032.g001
Introduction of the reserve decreases the yield at ΕMSY and by definition there is no alternative effort that produces the same maximal yet sustainable yield. The point of intersection in the bottom panel corresponds to a level of effort Ε1, where the yield is the same with or without a reserve. At levels of effort above Ε1 , the introduction of a reserve increases yield. In this scenario Ε1, is 150% of ΕMSY, so a fishery would have to have 50% excess effort for the reserve to be beneficial in terms of the yield of the target species. At even higher levels of effort (>150% ΕMSY ) the MPA mitigates the impact of overfishing and permits sustainable (but substantially reduced) yield.
The level of excess effort at which a reserve has a neutral impact on fisheries yield depends only on the ratio of movement out of the reserve (μ) to the maximum growth rate (r) (equation (15)). This relationship is shown in Figure 2a, when the movement rate is high relative to the growth rate, a reserve is beneficial at low levels of excess effort. The extreme situation where μ/r approaches infinity corresponds for example to a miniscule reserve, which clearly will have negligible impact on a fishery. At the other extreme,μ/r=0, there is no movement out of the reserve, consequently it will always have a negative impact.
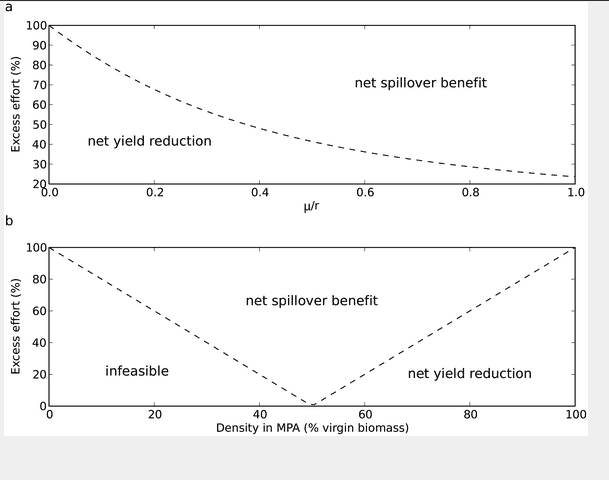
(a) The excess effort required for a reserve to improve fishery yield. For our simple model this was found to depend only on the ratio of the movement rate out of the reserve (and thus on reserve size) to the growth rate of the stock (). (b) The excess effort required for optimality as a function of the reserve density (at equilibrium). For example a reserve with 80% virgin biomass at equilibrium will provide a net economic benefit for a fishery that has more than 60% excess effort relative to optimal management. Combinations of excess effort and reserve density that fall in the bottom left region are infeasible; in these situations a reserve would have to decrease in population density after being formed (not possible in our model). Inside the “V” the reserve provides a net increase in fishery yield. In the right region the reserve decreases yield.
doi:10.1371/journal.pone.0107032.g002
Alternatively we consider the excess effort required for a reserve to be beneficial as a function of the reserve density at equilibrium (Figure 2b). If the reserve is at 50% virgin biomass density it has neutral effect on the fishery. This is because 50% virgin biomass corresponds to MSY in this model and all surplus production is moved to the main population through spillover. At reserve densities above this, a fishery must have more excess effort to benefit from a reserve. In particular if reserves have a high percentage of virgin biomass (a common conservation goal for reserves) they will only benefit fisheries that have greater mismanagement. For example, at 80% virgin biomass a reserve will only benefit fisheries with more than 60% excess effort.
Discussion
Model outcomes
The model presented here examines the circumstances under which spillover from a reserve is sufficient to increase fishery yield (thus providing a net spillover benefit). As expected, density of exploited species was higher in the reserve than the fished area, which may be mistaken in itself as evidence that the reserve will create a net beneficial increase through larvae production. However, it is important to consider the net effect, which in our model case was a decline in average density and a loss of yield except where effort exceeded ΕMSY. While models are by necessity a simplification of ecological complexity, we show that the extent to which effort must exceed ΕMSY for any yield benefit to occur from the reserve depends on the ratio of the rate of movement out of the reserve and the growth rate of the species concerned. Highly mobile/slow growing species received relatively less benefit from reserves where effort was above management targets compared to species with low movement/fast growth.
Our model is a relatively simple one chosen to illustrate a fundamental principle that is applicable across a broad range of fisheries. Different formulations for the biological model,ƒ(Nt) , can be specified and similar results were obtained for the Ricker model (not shown here). Three major assumptions were made to maintain model simplicity: spatial homogeneity, density dependence and steady state dynamics.
Spatial homogeneity is an inappropriate assumption for some species. For example, where there are clear source-sink relationships protecting the source in a reserve is likely to provide an overall benefit. The location of source areas can be consistent across different species and trophic levels, and in rare cases where these locations are known, it becomes possible to locate reserves that provide benefit to numerous, and theoretically all, species.
Density dependence in our model is a function of the total biomass in the local area (i.e. the fished population or the reserve population). This does not adequately capture the dynamics of species where density dependence varies substantially with age (e.g. density dependence occurring primarily during larval stages) and where different age classes have different movement rates across the reserve boundary. In such situations it could be possible for the reserve to provide a greater benefit by providing a recruitment increase to the fished region.
Steady state dynamics are widely used to explore fundamental fisheries principles. In the context of reserves, some models have shown that biological stochasticity may lead to theoretical net spillover benefits in fisheries where the biomass can be determined accurately on an annual basis and corresponding perfect catch limits set each year. Given the unrealistic nature of this assumption for most management situations there would be some value in further research that explored reserve benefits in a stochastic setting with realistic management. After the introduction of a reserve, it will take some time for the reserve population to build to the final density. Consequently it is expected that the reduction in yield will initially be much greater than predicted by our steady state model. With the concentration of effort the fished population would initially decrease before increasing some time later due to spillover from the reserve.
Our model did not consider that the introduction of a reserve may result in an effort reduction due, for example, to decreased accessibility or increased fishing costs. This would be beneficial for stock status and overall production in over-exploited fisheries, however, it would result in a reduction of production in well-managed fisheries.
Under our model there were no combinations of growth rate or movement where a net spillover benefit from reserves could occur unless effort exceeded ΕMSY . Where effort is less than ΕMYS, a loss of yield always occurs when reserves are implemented. The level of excess effort beyond ΕMSY at which a reserve provides net spillover benefits was shown to depend only on the ratio of movement out of the reserve to the rate of growth of the population (μ/r). We also showed that reserve configurations that achieve higher densities of stock are only beneficial for mismanaged fisheries (Figure 2b). For example, a reserve that ultimately increases biomass density to 75% of unfished levels would benefit a fishery if the initial effort exceeds ΕMSY by more than 50%. These results show that reserves will generally negatively impact yield for well managed fisheries. However reserves could minimize their impact on a well managed fishery by reducing the density increase of the fishery’s target species in the reserve. For example, a reserve could be of a sufficient size to protect species with small home ranges whilst being small enough that individuals of the target species frequently move beyond reserve boundaries (a high movement rate, ). This could also be achieved by having high reserve boundary length to total area ratios. The feasibility of this outcome will depend on the movement characteristics of the species involved.
Our finding that reserves cannot improve the yield of a well-managed fishery is consistent with several other theoretical studies [18]–[20]. The work here extends these findings by exploring the extent to which a fishery must be mismanaged before introduction of a reserve provides a benefit to the fishery in terms of yield.
Many fisheries have management objectives that constrain catch below the target of MSY assumed here, for example to manage risk from stochastic processes such as recruitment, or where there is an objective to target a maximum economic yield (MEY) that is variant to MSY. In these fisheries, effort and catch are lower than would occur with the MSY target, which reduces the negative impact of reserves on total yield, but also shifts the fishery further away from the level of depletion required for a net spillover benefit to occur.
Empirical context
The results from this study are consistent with other studies that have modeled the impact and/or benefits of reserves on fisheries in terms of improvements in yield. Following the publication of the early models on the potential net spillover benefits from reserves there have been surprisingly few empirical studies that have attempted to demonstrate the effect. Most of the reserve literature has concentrated on the changes within reserves, showing an increase in size and abundance of resident fish and crustaceans, particularly of reef associated species. Despite the lack of empirical evidence the argument persists that reserves will confer a net spillover benefit to fisheries. This view is actively promoted by government agencies. However, the literature confirms that the evidence for such a benefit is far from conclusive. Several studies report a lack of evidence for spillover due to the low movement at the scale of the reserve, while others showed that spillover occurred but not that lost yield was compensated to produce a net benefit.
While density dependent export from reserves is considered to be a rational expectation, no studies have been able to conclusively demonstrate a net spillover benefit, and leakage from reserves is probably more related to random movement within species. Several studies fail to provide conclusive evidence for net spillover benefits, yet argue that reserves are needed to provide fishery benefits. Spillover has been inferred from observations of a density gradient between the reserve and adjacent fished area even though evidence was acknowledged to be equivocal and where confounding factors such as a change in fishing practices or changed fisheries management strategies over the study period were ignored. Few of these studies consider whether the purported spillover to the fishery (as inferred from catch rates) has actually resulted in a net spillover benefit for the fishery. Even if CPUE goes up in a fished area it may be insufficient to result in a net production gain for the whole of the fishery.
Several studies have been able to demonstrate that spillover has contributed to an improvement in biomass and thus catch rate adjacent to the reserve . These examples, all in the Mediterranean, were conducted in areas where the total fishery had been severely depleted. In this respect they are similar to several studies in other areas that, on multiple lines of evidence, infer a net spillover benefit to fisheries. Examples come from Africa and Asia where the fisheries in question were over-exploited and where there was limited application and/or enforcement of standard fisheries management controls. The result was that the proclamation of a reserve resulted in a recovery of the population in the reserve and a subsequent improvement in catches close to the reserve boundary. This is consistent with our conclusion that reserves can provide a net spillover benefit for severely depleted stocks. It does not, however, provide evidence that the declaration of the reserve was the most efficient means of achieving that benefit.
There are many possible variations on the biological assumptions made in our model. Aspects such as stock heterogeneity and variant density dependence assumptions will influence the impacts of a reserve as well as the level of mismanagement, where a reserve switches from being beneficial to being detrimental for a fishery.
The model results presented here are for a general case, which is appropriate for consideration of reserves where a large number of species with variable life histories and spatial distributions are affected by change in management. Closed areas for traditional fishery management purposes are applied on a species by species basis and may have very different management outcomes to reserves because they can be designed and located to affect an individual stock. There are numerous cases where species with spatial heterogeneity, such as spawning aggregations or larval source-sink dynamics, benefit from fishery closures that target important source areas. A total fishing closure would achieve the same result for those species, but can be expected to have less beneficial results for other exploited species.
Conclusions
We conclude that in fisheries where there is effective management, marine reserves are unlikely to produce a net spillover benefit for the total fishery, whereas they may be beneficial where the fishery has been mismanaged and stocks severely depleted. These results expand the implications of previous work by providing estimation and evaluation of the degree of mismanagement of fisheries that is necessary for non-specific closures to provide net benefits to fisheries.
The conclusions from the modeling presented here are supported by review of empirical studies, where spillover benefits have only been conclusively demonstrated in highly depleted areas. Together with the combined weight of earlier modeling work, they suggest that a net benefit from spillover should not be expected in areas already benefiting from quality traditional fisheries management.
These generalised findings in relation to reserves should not be confused with the use of targeted spatial closures for single fisheries, where it is possible to increase yield through closures by taking account of the spatial heterogeneity of life history traits.
While reserves may be proclaimed for a range of conservation objectives (including addressing impacts such as the effect of fishing on benthic environments, interactions with threatened species and catch of non-target species), we contend that it is misleading for governments to promote reserves on the basis of net spillover benefit in the context of well-managed fisheries. Reserves are only likely to be an effective strategy for fisheries management where effort is not or cannot be effectively controlled across the wider stock.
November 2014
