
© C Greg Lutz
Some 150 years ago, global poultry production looked quite a bit like tilapia production did in 1970. Chicken production was seasonal in many parts of the world because of climate and nutritional constraints, and it was a very fragmented industry, focusing mostly on local or regional markets. As a result, there were hundreds of localised chicken breeds and the culture of different birds in different localities was the norm.
Throughout the world the past century saw a shift from this fragmented industry, based on subsistence- and small-scale production, to vertically integrated facilities and specialised hatchery businesses with a focus on genetic improvement. This trend continues today, as industrial production expands in less developed nations.
The original focus of chicken breeders involved both egg and meat production in the same birds, but this shifted to separate breeding goals after World War I. At that point, production of meaty birds, known as broilers, became distinctly differentiated from that of the layers that supplied eggs. When broilers were first commercialised, mass selection was the approach used to improve production and meat yield. This resulted in moderate progress, because at the time most breeding flocks exhibited heritabilities ranging from 0.2 to 0.4.
In the tilapia industry the importance of meat yield is finally gaining more recognition as an important breeding goal. Heritabilities for fillet yield, ranging from 0.2 to 0.38 from 130 to 210 days of age, were reported in a population of Nile tilapia in Brazil (Turra et al. 2012) and a heritability of 0.25 was reported from Malaysia (Nguyen et al. 2010), but Gjerde et al. (2012) reported a much lower value of 0.06 for a line from Nicaragua. More recently, Yoshida et al. (2019) reported a modest increase in heritability estimates when using genomic information on a population in Costa Rica.
In chickens we see a negative correlation between growth and fecundity, so reproductive capacity is important in female lines, even when meat production is the ultimate goal. In tilapia improvement programmes, growth is also usually the breeding goal, but spawning success and frequency are critical considerations in commercial hatcheries. Heritability of spawning success was reported as 0.14 to 0.22 for a population of Nile tilapia in Vietnam, depending on the statistical model and spawning period.
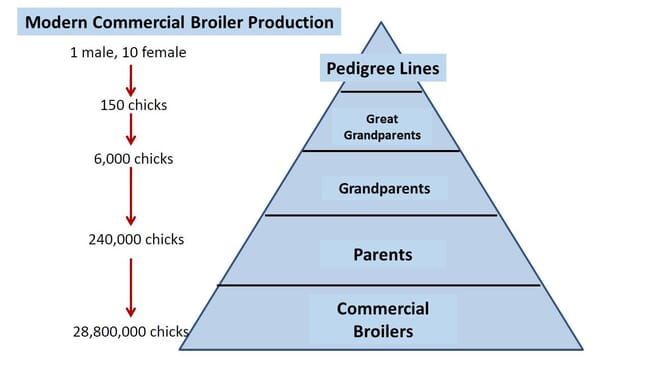
In the years following World War II, chicken breeders began to cross distinct lines to produce broiler chicks. One reason was to maintain “female” lines that exhibited good egg production while crossing these with male lines that might be less productive as a result of intense selection for growth. This same practice occurs in many tilapia hatcheries around the world. Modern broiler production has evolved into a five-year process where four distinct lines are crossed to ultimately produce chicks. Crossing distinct lines is still not a common practice in tilapia production, but for a period of time hybrids were quite popular in China and also in Israel. At one time the breeding programme of a large tilapia operation in Honduras was based on a double crossing design similar to those used in the chicken industry.
Disease resistance has been an important component of chicken breeding programmes for many decades. In tilapia, a similar emphasis is taking hold. The recent emergence of tilapia lake virus (TiLV) has caused concern across the globe, but new research suggests breeding programmes to reduce or eliminate this threat may be possible. Using data from 124 O. niloticus families in Malaysia, researchers estimated heritabilities for TiLV resistance ranging from 0.48 to 0.56, with family survival ranging from 0 percent to 100 percent (Barria et al. 2020).
Some of the more long-standing tilapia pathogens are still causing major problems. Many tilapia hatcheries struggle with Flavobacterium columnare, the causative agent for columnaris disease. Selective breeding has been shown as a means to improve resistance to columnaris in tilapia, and this is important in tropical regions, where vaccines are not available and antibiotic resistance is on the rise. Researchers working with the Chitralada 4 strain in Thailand reported heritability estimates for columnaris resistance ranging from 0.14 to 0.30, based on different statistical models, with family survival rates ranging from 8 percent to 70 percent (Wonmongkol et al. 2017). Genetic influence over resistance to Streptococcus iniae and S. agalactiae have also been demonstrated, with heritabilities of 0.52 and 0.38, respectively, but resistance to one pathogen does not appear to confer resistance to the other (Shoemaker et al. 2016). Dr Carlos Lozano, with Spring Genetics, stated that “selection for disease resistance” is among the tilapia industry’s top priorities for genetic improvement, adding “In particular diseases like S. Iniae, S. agalactiae, Francisella,
TiLV and columnaris.”

© C Greg Lutz
By the 1960s, chicken improvement programmes were emphasising the importance of feed conversion efficiency. This trait is quite difficult to measure in fish, but recent study demonstrated significant genetic control over it in Nile tilapia, with an estimated heritability of 0.32 (de Verdal et al. 2018). One genetic correlation with feed efficiency that was significant was weight loss at fasting (0.80), which would allow indirect selection for individuals with superior FCR while avoiding the complex task of measuring individual feed intake.
As quantitative genetic tools and selection programmes became more sophisticated, the chicken breeding sector began to consolidate, and most breeding programmes adopted selection indices that included more than 40 distinct physiological traits relating to feed efficiency, disease resistance, skeletal structure and behaviour. By the late 2000s only three major breeding companies dominated the market: Cobb-Vantress, Aviagen and Group Grimaud (a multi-species company based in Europe that also has interests in aquaculture species).
Over time, genetic improvement tools for chickens have “evolved” from mass selection, to index selection, to the use of BLUP estimates, to marker-assisted selection and now genomic selection is becoming the norm. When I asked Dr Lozano to cite some of the most important developments in tilapia improvement in recent years, he indicated: “The recent development of SNP chips for tilapia has enabled the inclusion of marker assisted (MAS) and genomic selection in tilapia breeding programmes (eg Spring Genetics and GenoMar). This is especially important for disease resistance, where generally family selection was used and candidates were selected based on the performance of their siblings. With genomic selection we can select candidates based on their individual genetic merit. Gene-editing technologies have also recently been applied by AquaBounty and approved for Nile tilapia in Argentina.”
Since there are so few genetic nuclei, modern industrial chicken lines must be adaptable to conditions found around the globe. Unlike a century ago, when any number of local chicken breeds could be found around the world, the birds being produced by these large corporations must incorporate adaptability to a variety of grow-out conditions and diets. This potential conflict between centralised breeding operations and a multitude of grow-out environments may eventually become an issue for tilapia producers as improved lines are disseminated across the globe. When asked about this, Dr Lozano commented: “I envision the dissemination of superior genetics mostly to industrial scale producers. For small scale producers the low profit margin may restrict their accessibility to high quality fry.”
Modern chickens are largely descended from the red jungle fowl, but at least one other related species appears to have contributed to the birds we raise today. Many lines of O. niloticus also include genetic contributions from other species. However, while the genome of the modern chicken has been relatively stable over the past century, this milestone is yet to be attained in tilapia. Not all tilapia are created equal. Researchers in Thailand used microsatellite genetic markers to examine changes in four lines of Nile tilapia (Oreochromis niloticus) derived from the genetically improved farmed tilapia (GIFT) strain, and in two lines derived from a local Chitralada strain. Three of the four GIFT-derived populations remained relatively “pure”, with no decline in genetic variation. However, one of the GIFT-derived populations showed high levels of introgression from the Chitralada strain, and the Chitralada-derived populations exhibited introgression from GIFT lines. Introgression from O. mossambicus was also observed in one of the Chitralada-derived lines and from O. aureus in one of the GIFT-derived lines (Sukmanomon et al. 2011).

© C Espejo Genipez Viterbo
The situation regarding some semblance of genomic standardisation becomes even more complicated with red tilapia. In many parts of the world, consumers demand red-coloured fish. Although red colouration has been established in O. niloticus (the “red Stirling” line) through the isolation and propagation of a single red mutant (McAndrew et al. 1988, Hussain 1994), most strains of red tilapia include genetic contributions from several distinct species including slower-growing O. mossambicus and O. urolepis hornorum (Desprez et al. 2006). In recent years efforts to develop robust commercial lines of red O niloticus have advanced through backcrossing red Stirling fish with fast-growing strains such as GIFT and Chitralada (Lago et al. 2017). In Colombia, my colleague Carlos Espejo has adapted a similar strategy, crossing local red lines of multi-species ancestry with Chitralada-based wild-type fish.
Just as most “unimproved” varieties of chickens would not perform very well in commercial chicken houses, most industrial birds would not survive long in the farmyards of subsistence farmers around the globe. Many local varieties of chicken persist around the world, and leaving aside all the introgression and hybridisation that has occurred over the past century, even within pure O. niloticus a similar array of variation can be found in terms of local and regional strains. Africa as a whole is a region of concern in terms of conserving genetic resources scattered among hundreds of local stocks of Nile tilapia. In the case of chickens, many historically recognised European, Asian and North American breeds were already reported as extinct by the FAO in 2007, having been replaced by modern varieties. Many more breeds were listed as at risk at that time. However, in regions where subsistence farming still plays a significant role in food production, indigenous chicken breeds persist, due to their accumulated adaptation to local conditions. Hopefully the same will hold true for tilapia.
References available from the author.





