In intensive animal production, high daily weight gain and high feed efficiency are essential. However, high performance is associated with increased levels of stress.
Stress – such as from high stocking density, pathogenic pressure and temperature – is associated with enhanced levels of reactive oxygen species (ROS) and linked to sub-optimal antioxidant status. Selenium (Se), in this respect, is a very important essential trace element as it is a vital component of selenoenzymes (eg glutathione peroxidase, GPx) which play a role in reducing ROS and to maintain a healthy antioxidant status. A disruption of this steady state causes tissue damage due to interaction of ROS with lipids, proteins and DNA. These negative interactions reduce their metabolic activity.

In order to maintain this steady state a continuous as well as optimal selenium supply is essential. However, this can be difficult to achieve when uptake from the diet is impaired when stress is present. At that moment selenium is in high demand, to produce selenoenzymes and combat ROS. Selenium storage inside the animal, in that respect, would be beneficial. This article provides an overview of the scientific literature on the beneficial effects seen with the addition of L-selenomethionine to the diet and this focussed on salmon, trout and tilapia. Results from a recent trial on tilapia conducted in Thailand are discussed.
Maintaining an optimal selenium steady state: a nutritional solution
Selenium can be added to the diet in either inorganic or organic forms (Figure 1). The advantage of using organic selenium (L-selenomethionine, L-SeMet) over inorganic sources (eg sodium selenite or selenate) is its ability to be incorporated directly, without conversion, into general body proteins as a methionine source. L-selenomethionine is the only selenium compound that has this ability. The incorporated selenium, in the form of L-selenomethionine, acts as a storage of selenium in the animal. This stored selenium ensures optimal supply, even during stressful periods.
If necessary, the stored selenium gets metabolised to selenide (H2Se) then to de novo
selenocysteine (SeCys). This molecule will be incorporated, as the active site, in selenoproteins. Other selenium compounds, such as SeCys and sodium selenite, are not storable but will be metabolised to de novo SeCys. These compounds will be quickly excreted when intake is in excess. L-selenomethionine will only be metabolised to selenide when there is a need. This form is therefore less prone to excretion and toxicity reactions (Rayman, 2004).
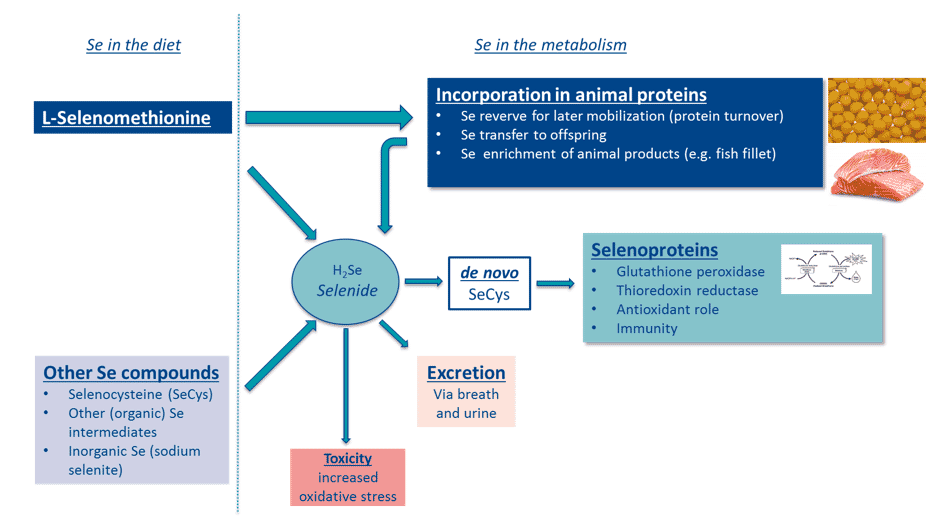
The metabolism of L-selenomethionine and other selenium compounds © Adapted from Rayman, 2004; and Combs, 2001
Aquatic protein challenge: a case for L-selenomethionine
Traditionally, fishmeal was the preferred protein sources in aquatic feeds. Due to limited availability, pressure on wild fish stocks and variable prices there is an interest in alternative, sustainable protein sources. Plant meals, for example, are suitable alternatives in the growing global aquaculture industry. However, replacing marine ingredients in fish feed with plant sources changes the nutrient composition of the feed. Selenium concentration of fillets is reported to be highly impacted by high levels of substitution, reducing the added value of fish consumption (Lundebye et al. 2017; Betancor et al. 2016).
Although selenium levels are decreasing within the fish, the demand for selenium to protect against (oxidative) stress remains. Stressors (eg environmental, metabolic) are an important issue for the productivity and profitability of fish farms. These stressors may cause increased oxidative damage to lipids, proteins and DNA and increased mineral mobilisation from tissues and their subsequent excretion. High stress may therefore lead to increased mineral requirement. L-selenomethionine is established to be a highly available selenium source leading to higher selenium deposition compared to inorganic selenium sources (Figure 2). It can therefore counteract selenium depletion caused by plant-based diets.
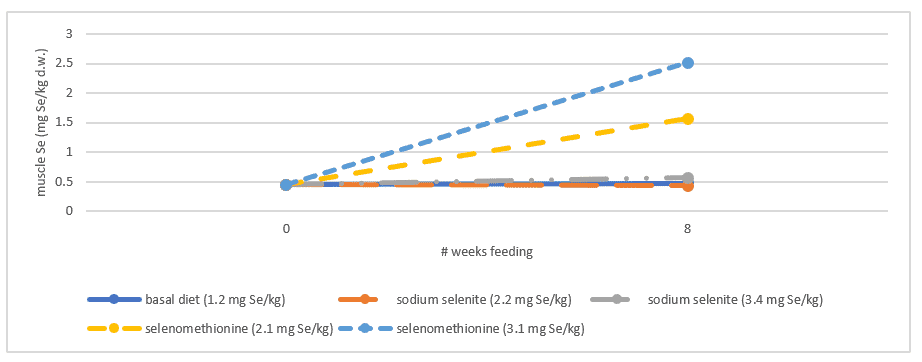
Selenium concentrations in muscle (mg Se/kg dry weight) of Atlantic salmon fed a fishmeal-based diet supplemented with sodium selenite or selenomethionine at levels of 1 and 2 mg Se/kg feed, respectively, for 8 weeks. © Lorentzen et al, 1993
Control stress and win
Dietary selenomethionine supplementation is known to offer a way to reduce performance loss under stress, such as crowding conditions (Küçükbay et al. 2008). A recent study, performed at the Mahasarakham University, Thailand, showed increased performance and high protection against pathogenic pressure. A total of 735 Nile tilapia (initial weight 13.52±0.5g) were fed one of seven experimental diets (in triplicate) in fibreglass tanks for eight weeks. Organic Se (L-selenomethionine, SeMet; Excential Selenium 4000, Orffa Additives BV) and inorganic Se (sodium selenite, Na2SeO3) were each added to the basal diet at 1, 3, and 5mg Se/kg. The basal diet (28 percent crude protein), without Se supplementation, was used as a control.
The final Se concentration of the basal diet was 0.68mg Se/kg. Organic and inorganic Se supplemented diets contained 1.78, 3.53 and 4.90mg Se/kg and 1.75, 3.49 and 5.30mg Se/kg, respectively. Fish were fed at 5.0 percent of their body weight twice a day. Parameters were assessed at the end of the rearing period. After eight weeks, 20 fish from each treatment were challenged with intraperitoneal injection of the virulent Streptococcus agalactiae serotype III at 1x107 CFU/mL. The cumulative mortality was observed for 21 days and the relative percent survival (RPS) was calculated.
Table 1 shows that weight gain (WG) of fish fed SeMet at 1mg Se/kg was significantly higher than that of fish fed basal diet (p<0.05). Lymphocytes were significantly (p<0.05) higher in fish fed SeMet (1mg Se/kg) compared to fish fed basal diet. Alanine transaminase (ALT), aspartate transaminase (AST), creatinine, blood urea nitrogen (BUN), albumin, globulin and total protein were not significantly influenced by dietary Se supplementation.
Increasing dietary Se level, particularly in the form of SeMet, led to a decrease in serum cholesterol concentrations. Interestingly, the innate immune response (eg lysozyme, catalase, myeloperoxidase, superoxide dismutase and glutathione peroxidase) activity was significantly (p<0.05) increased with Se supplementation compared to the basal diet group, especially for fish fed SeMet (1 and 3mg/Se kg). Malondialdehyde (MDA) in fish serum, on the other hand, was decreased numerically for all supplementation levels. Fish fed SeMet (1mg Se/kg) showed the highest RPS after the challenge with S. agalactiae.

Growth performance, haematological values and immune parameters of fish fed experimental diets. Note: Values show mean, pooled SEM, n = 90; S= L-SeMet (Excential Selenium 4000) and sodium selenite (Na2SeO3), L=level of selenium supplementation. Values in the same row with different letters differ significantly (p < 0.05). White blood cells (WBC), alanine transaminase (ALT), aspartate transaminase (AST), blood urea nitrogen (BUN), malondialdehyde (MDA), relative percent survival (RPS).
Conclusions
L-selenomethionine (Excential Selenium 4000) was tested and validated by independent researchers around the world in peer-reviewed publications (eg Berntssen et al. 2018; Silva et al. 2019) and proven to be effective in increasing the selenium and antioxidant status of fish, even under challenging conditions. This will result in improved performance and immune function. Very high levels of L-selenomethionine (5 mg Se/kg feed) do not appear to have negative effects on performance nor on immune parameters.
L-selenomethionine therefore has a good application in fish diets when fish are kept under stressful conditions or in any diets where fishmeal is replaced by plant meals. L-selenomethionine helps to maintain selenium concentration in fish fillets and therefore contributes to the positive healthy image of fish consumption for humans.
References available, on request, from bruneel@orffa.com


