Here, Fredrik Jutfelt et al, University of Gothenburg, Sweden, show several severe behavioural disturbances, including effects on boldness, exploratory behaviour, lateralisation, and learning in a temperate fish, the three-spined stickleback (Gasterosteus aculeatus). The behavioural effects were consistent throughout the exposure period and increased in effect size with exposure time. We observed the effects on adult sticklebacks, a species known to be tolerant to other environmental stressors. Our findings suggest that behavioural abnormalities that stem from CO2 exposure are not restricted to sensitive tropical species or early life stages and may therefore affect fish on a global scale. The severity of disturbances and the possibility of a serious behavioural problem for fish across the globe is cause for concern.
Introduction
Anthropogenic CO2 emissions are increasing the atmospheric CO2 concentration, which drives increasing dissolution of CO2 into the oceans through a process commonly referred to as ocean acidification. Teleost fish have been regarded as highly tolerant to the effects of ocean acidification. As fish are active organisms with a high metabolism, their muscle activity produces CO2 at a variable rate. Consequently, they experience internal fluctuations in CO2 concentration. The gill is the major organ for pH regulation where hydrogen ions are excreted and bicarbonate ions absorbed. These mechanisms effectively buffer the blood pH during transient periods of high water CO2 concentration, and thus, fish have been assumed to be tolerant to the relatively modest CO2-challenge of ocean acidification. Maintaining blood pH despite CO2 exposure, however, requires modifications in concentrations of blood ions, including increased HCO3-, decreased Cl-, and increased Na+ concentrations.
Recently a series of studies from an Australian group reported unexpected behavioural effects in several coral reef species. The effects include olfactory disturbance and olfaction preference reversal in larval clownfish; impairment of visual risk assessment in juvenile damselfish (Pomacentrus amboinensis); reduced auditory response and learning in clownfish (Amphiprion percula); reduced olfactory prey detection, reduced feeding, and increased activity in brown dottyback (Pseudochromis fuscus); and reduced lateralisation and prey detection in a larval Neopomacentrus azysron, another coral reef species. Atlantic cod larvae were found to be behaviourally resistant to very high pCO2 (4200 matm), indicating that sensitivity to CO2 changes can be highly variable among teleost fish. It has been suggested that fish living at the warm edge of their tolerance range are more sensitive to CO2-induced acidification. As a result, tropical reef fish have been suggested to be among the most sensitive.
Nilsson (2012) suggested a neural mechanism as the cause of some or all of the observed behavioural effects of CO2 exposure. As fish experience increased CO2 with accompanying acidosis, they partially reduce the acidosis via the Cl2/HCO3- exchanger in the gill epithelium. This leads to decreased Cl- concentration as well as increased HCO3- concentration in the extracellular fluid. The GABAA receptor is a major vertebrate inhibitory receptor, and its main mechanism is hyperpolarisation of the postsynaptic neuron by the function as an ion channel permitting the passage of negatively charged Cl- and HCO3- ions. Reduced extracellular Cl- concentrations reduce the electrochemical gradient for Cl- influx, while increased intracellular HCO3- concentrations may lead to efflux of anions and cause depolarisation of post-synaptic neurons and therefore possibly a reversal of normal GABAA receptor function. The ubiquitous nature of this receptor in the vertebrate brain suggests that this mechanism likely affects many aspects of behaviour. To date, however, only one study has tested this hypothesis.
The three-spined stickleback is a species with remarkable physiological plasticity and high tolerance to fluctuations in water chemistry and temperature. This species is found in a wide range of habitats, from marine to limnic and Arctic to subtropical waters. The ability to acclimatise to such wide environmental conditions suggests that these fish, in theory, may be tolerant to the increases in pCO2 predicted for the end of the century.
Thus, to test the hypothesis that temperate fish are more tolerant to CO2 exposure than tropical reef fish we exposed sticklebacks to control water (330 μatm CO2) or high pCO2 water (991 μatm CO2) to mimic present-day surface CO2 concentrations and a predicted scenario for CO2 concentrations in the year 2100 (the fossil fuel intensive IPCC A1F1 emission scenario [20]), respectively. A range of behavioural tests were performed after 20 days of exposure and repeated after 40 days of exposure to test for long-term acclimation capacity. The behavioural tests were lateralisation, a measure of the behavioural asymmetry of individuals; novel object test, a widely used method for assessing boldness and curiosity in fish; and an escape chamber test designed for testing exploratory behaviour and boldness.
Materials and Methods
Ethics statement
Rearing, handling and experimental procedures were approved by the ethical committee on animal experiments of Gothenburg, Sweden (ethical permit: Fredrik Jutfelt 100-2010 and 151-2011).
Experimental animals
The study was conducted at Sven Love´n Centre for Marine Sciences, University of Gothenburg, Kristineberg (Sweden) in July and August of 2012. Marine female sticklebacks (Gasterosteus aculeatus) were caught using a seine net in Sa¨ lvik, Fiskeba¨ ckskil bay, Lysekil, Sweden (geographic coordinates: 58u149500N11u279 300O). A total of 100 fishes were distributed into ten 25 L glass aquaria (ten fish each). Five aquaria were randomly assigned "control" and five "CO2". The aquaria were constantly supplied with flow-through seawater at a rate of two L min-1 from one of four header tanks (200 L). Each header tank had a flow of five L min-1 of flow-through seawater taken from five m depth, and constant aeration. The fish were kept at 14 h:10 h light:dark cycle and fed ad lib twice daily with frozen Artemia naupli. Water salinity, oxygen saturation, temperature and pCO2 were measured daily and alkalinity was measured weekly. Oxygen saturation remained above 90 per cent in all measurements. The water temperature was 17.6-°C±1.2 (SD) and salinity averaged 24.2 PSU±3.4 (SD).
Experimental treatments
The pCO2 of all aquaria was measured daily using direct pCO2 measurements with an infra-red CO2 probe (GMT 222, Vaisala, Finland) connected to a submerged gas-permeable silicone membrane and the air inside the membrane was circulated in a closed loop to equilibrate with dissolved pCO2, as described elsewhere. Correct factory calibration of the probe was confirmed on several occasions during the experiment using water thoroughly bubbled with a gas mixture of 1010±10 ppm CO2 in air (AGA, Sweden).
The pCO2 of the two CO2 treatment header tanks was maintained at the target value of 1000 matm using pH stat Computers (Aqua Medic, Bissendorf, Germany) connected to solenoid valves regulating administration of 100 per cent CO2 gas (AGA, Sweden). The pCO2 of control aquaria was 333 μatm±30 SD and the pCO2 of the CO2 treatment aquaria was 991 μatm (57±SD). The variance in pCO2 between aquaria within each header tank system was below the detection limit (
Half of the fishes were exposed to increased pCO2 over a period of 43 days while the other half was kept in control water. Aquaria were randomly attributed control or elevated CO2. The fish were allowed to acclimatize to the aquaria for ten days before the CO2 exposure started.
Behaviour experiments
The behavioural experiments were carried out in the same environmental conditions (temperature, salinity, light, pCO2) as during the exposure. 20 fishes from each treatment (randomly netted) were used in each experiment. All tests were done by direct visual observation. Experiments were performed using individual sticklebacks gently introduced into the experimental chambers, always at the same starting point and position. PlastKapTek, Sweden manufactured the acrylic experimental chambers. The behavioural tests were performed twice, on four consecutive days each time, with one behavioural test each day. The tests were performed on days 20–23 and 40–43 after initiation of exposure.
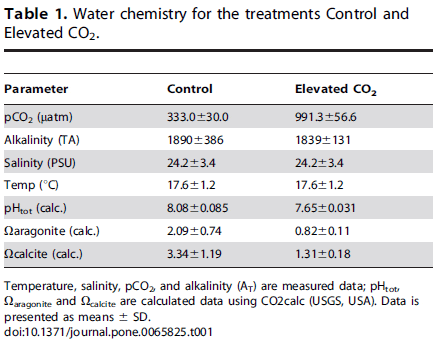
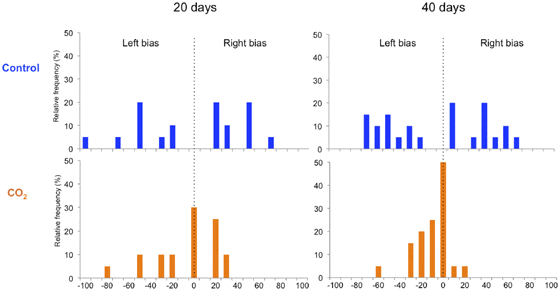
Lateralization. Fishes were individually introduced into a double T-chamber (dimensions: 50 cm long with a 9 cm wide double T-channel (figure 1), according to Domenici et al. 2012) and gently encouraged by a plastic rod to move forward until a left or right turning choice was made. The procedure was repeated twelve times for each of 20 fish at day twenty, and twenty times for each of 25 fish at day 40. The purpose of increasing the number of turning events and n at day 40 was to reduce variance and increase statistical power. The relative and absolute lateralization indexes were calculated according to Domenici et al. 2012.
Novel object. Fishes were left to settle individually in a test aquarium for 3 min, followed by the introduction of the novel object (a 5 cm Rubik’s cube) to the aquarium. The time each fish spent investigating the object was recorded during 7 minutes. The novel object test is intended to measure fear of novelty, and can thus be used to categorize fish on a bold to shy axis.
Escape challenge. Fish were placed in a white vertical cylindrical chamber (Ø16 cm69 cm with a Ø5 cm exit hole on the side), with their heads facing away from the chamber’s exit. Time spent inside the chamber until escape was recorded. The procedure was performed twice for each fish. We had not seen this experiment described elsewhere and it was designed to assess exploratory behaviour after pilot studies indicated reduced exploratory behaviour.
Statistics
No tank effects were detected (one-way ANOVA, p>0.1), so tanks were pooled within treatment. Statistical differences between treatment groups were tested for using an independent samples ttest with equal variances not assumed in SPSS. Differences are considered statistically significant when p<0.05, and p-values are given in the figures. Data is presented as mean ± SEM unless otherwise noted.
Results
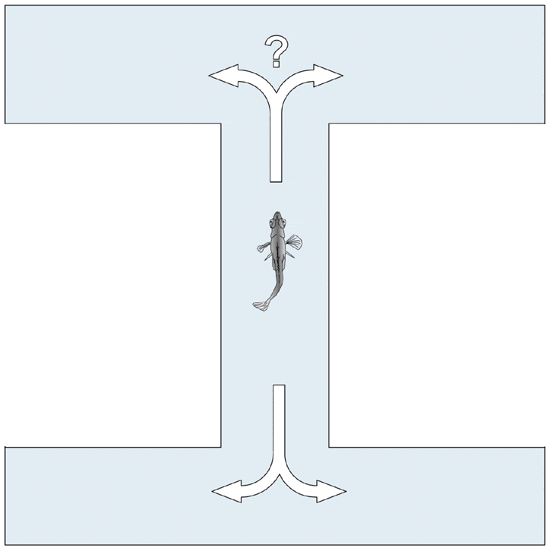
The fish gained weight during the experiment (Figure 2) and the treatment did not significantly affect final weight (p = 0.14). Total mortality over the whole experimental period was high as expected in post-spawning females: 36 per cent in the control group and 28 per cent in the CO2 group, but was not affected by CO2-exposure (p = 0.59).
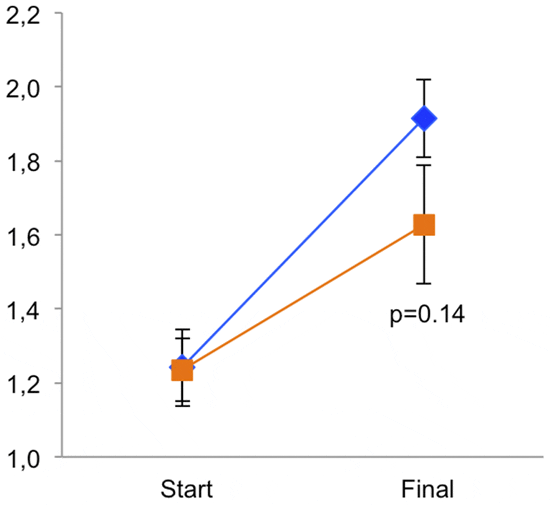
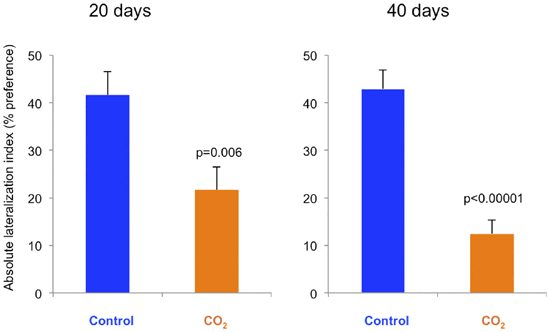
The control fish showed a wide spread of turning preferences, shown as relative lateralization index, while the CO2-exposed fish had a reduced distribution (figure 3). This difference is shown as absolute lateralization index (Figure 4) where the average turning preference is reduced by CO2-exposure. At day 20 the difference is a factor of 2 (p= 0.006), while at day 40 the effect size is larger with a factor of 3.6 (p<0.00001).
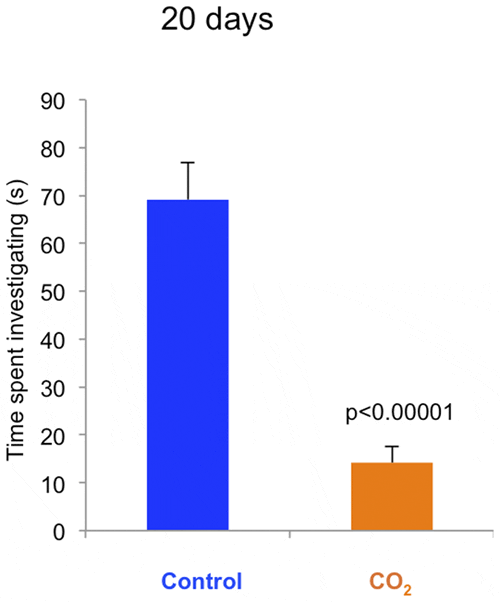
When presented with a novel object control fish showed curiosity and explored the object. The average time spent investigating the object was five times longer for control fish than CO2-exposed fish (p<0.00001) at day 20 (figure 5). On day 40, neither group showed interest in the Rubik’s cube, possibly because the fish had examined the object on the previous test occasion (data not shown).
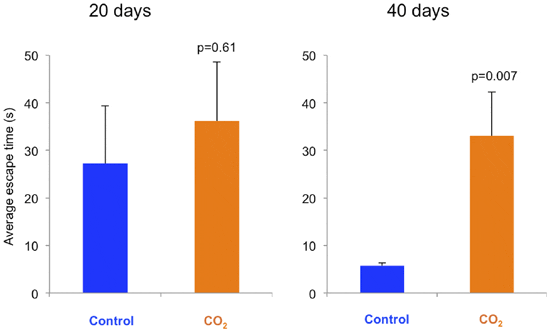
Both control fish and CO2-exposed fish took roughly half a minute to escape the escape chamber at day 20, with no significant effect of treatment (figure 6). At day 40 the CO2-exposed fish took the same amount of time to escape the chamber while the control fish were six times faster (p= 0.007).
Discussion
We found surprisingly strong effects in all of the behavioural tests performed. This indicates that most, if not all, of the sticklebacks’ behavioural repertoire can be affected by CO2.
As expected, the control fish turned to their preferred side 70 per cent of the time on average. In contrast, CO2-exposed fish lacked a preference and turned to each side 50 per cent of the time. A similar reduction in lateralisation was previously described in larvae of a coral reef fish exposed to CO2, indicating that this effect may be widespread both geographically as well as among species and life stages. A behavioural side preference is beneficial in a range of situations, such as multi-tasking, orientation, and escape. Any disturbance in behavioural lateralisation may therefore reduce fitness.
When fish were presented with a novel object in the form of a small Rubik’s cube on day 20 of the experiment, control fish examined the object five times longer than CO2-exposed fish. As the novel object test is a well-established method for estimating boldness and curiosity in fish, the finding likely indicate that CO2-exposed fish were less bold and/or less curious than control fish, with unknown fitness implications.
The difference in escape time at day 40 was mainly due to reduced escape time by the control group. The experiment was not specifically designed to test learning, yet the improvement by control fish in escape time from day 20 to day 40 is possibly due to memory of the escape technique from the previous trial. The CO2- exposed group showed no improvement at the second test, suggesting decreased learning ability. Learning is vital for acquiring new skills and behaviours. Interaction with predators is an efficient way of learning the identity of the predators, albeit sometimes with immediate costs (i.e., injury). Fish can learn to recognise predators through their own experience, through cues from injured conspecifics, and from social learning. Any impairment in learning ability may therefore affect fish survival through reduced predator avoidance.
Sticklebacks are physiologically plastic with a large potential for acclimation to environmental challenges, such as changes in salinity or temperature and environmental toxins. Acclimation to large variations in salinity and temperature commonly takes days to weeks in temperate fish. Additionally, G. aculeatus have short life spans of an average of 3 years; therefore, a 40-day exposure is considered long-term compared to the life span of sticklebacks and should be long enough to allow maximal acclimation. Because any possible acclimation failed to restore normal behaviours in the exposed fish, evolutionary selection may be required for sticklebacks to develop tolerance to high CO2 levels. The physiological reason behind the lack of acclimatory capacity is unknown; however, GABAA receptors may have be adapted to a narrow range of ambient CO2 levels, whereas at higher ambient pCO2, the receptors are unable to function correctly due to the associated modification of extracellular and intracellular ion concentrations.
The three behavioural tests employed in this study are artificial, and so the relevance of these tests to ecological fitness is largely unknown. In addition, the CO2 treatment had no significant effect on survival or growth over the 40-day exposure period, suggesting that the behavioural effects are not severe enough to affect performance in an aquarium setting with ad lib food supply. However, because performance on all three tests, and tentatively learning, was severely degraded, it is likely that the majority of the fishes behavioural repertoire was affected by CO2 exposure. From a physiological perspective, ubiquitous changes in behaviour are consistent with the proposed neurophysiological mechanism involving altered GABAA function, as GABAA receptors are omnipresent in the central nervous systems of vertebrates and any disturbance to the GABA system would affect most behaviours. Thus, complex behaviours, such as prey capture, predator avoidance, and mating rituals, would likely be disturbed in a natural setting with subsequent negative fitness consequences.
Conclusions
The current study provides indications that ocean acidification could adversely affect fish behaviour on a global scale this century. As sticklebacks are highly tolerant to many other environmental factors, it is unlikely that these animals are unusually sensitive to CO2 exposure. Severe behavioural effects have been documented in a few coral reef species (all from the order Perciformes) and here in temperate sticklebacks (order Gasterosteiformes). Thus, CO2 could potentially affect the behaviour of many, if not most, marine teleost species. As acclimation to counter the effects was not detected, despite the long exposure time, the most pressing issue now is the potential of fish to inheritably adapt to high CO2 levels to counter these behavioural disturbances. If the evolution of tolerance to CO2 is slower than the rate of CO2 increase, the ecological consequences of ocean acidification could be severe.
November 2013



