Introduction
Moulting is an inherent habit of crustaceans, in which periodically the old exoskeleton is cast off and a new one is formed. It allows further growth or weight increment. Moulting is “the continually recurring crisis in the life of the decapods crustaceans for it is both dangerous and expensive”. The moult cycle is the most critical and challenging phase in crustacean physiology. Moulting leads to the total replacement of old exoskeleton, including appendages, with a new exoskeleton. The shed exoskeleton is called the exuvium.
Detailed investigations were made on the moulting phenomenon of Macrobrachium rosenbergii. In giant freshwater prawns, moulting generally takes place at night and this cyclic process continues until death. In addition to growth, moulting also leads to varied changes in the general behaviour of the prawn. Since moulting affects the regular behaviour of crustaceans significantly, it deserves to be documented.
Moult cycle
 |
| Entire shed-off shell of a prawn |
Although moulting appears to take place as a single event, it is a multistage process. The moult cycle in M. rosenbergii has been defined as post-moult, inter-moult, pre-moult and moult. These stages might not be distinguished very clearly. But the behavioural differences during different stages can easily be observed.
The general physiological changes during different stages in a moult cycle are given below:
Post-moult: (Metecdysis/ Soft back stage)
This stage falls a few days after moulting. This stage is characterized by hardening of the newly formed exoskeleton by deposition of calcium and other minerals and results in the complete formation of a new exoskeleton.
Inter-moult: (Anecdysis)
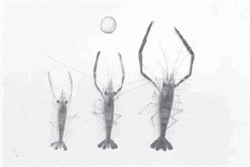 |
| Male morphotypes of prawn |
This is the stage between successive moults. The prawn in this stage is called an instar. The water taken up during ecdysis is gradually replaced by tissue growth, enrichment of organic matter and mineral reserves, resulting in growth of the prawn.
Pre-moult: (Proecdysis/ Peeler stage)
This is the stage just before moulting. In this stage, calcium is reabsorbed from the old integument leading to enlarged epidermal cells.
There will be clear separation of the old integument from the epidermis for moulting. Appendages also regenerate.
Moult: (Ecdysis)
The animal comes out of the old exoskeleton with a newly formed soft exoskeleton underneath and absorbs water in the tissue for increase in size. The above stages are broad classification of the moult cycle, which give some idea of the physiological process that takes place during moulting.
Moult cycle duration
In M. rosenbergii, the length of the moult cycle varies depending on the size, sex and age of the animal. However, it is not clear what is the exact duration of each stage. In Penaeids generally the moult cycle is short for juveniles and long for adults. Giant freshwater prawn exhibits long (about 30-80 days) inter-moult periods, short (about 10-12 days) pre-moult and very short (about 2-6 days) post-moult periods. In Penaeus merguiensis, actual moulting lasted for 40 seconds and in P. duorarum it is only 20-30 seconds.Changes in the body chemistry
Calcium levels fluctuate during the moult-cycle, indicating the importance of this mineral for exoskeleton formation. Crustaceans have different degrees of calcium demands that change depending on the stage of moulting. In freshwater prawns, as in other crustaceans, calcium supply is largely from the mobilization of calcium from the exoskeleton. In M. rosenbergii addition of vitamin D3 interferes with the metabolism of calcium and inorganic phosphorus in the tissues, and alters moulting frequency.
Steroid hormones control moulting, the precursor is ecdysone, an ecdysteroid. The steroid hormone concentration in the haemolymph fluctuates, being low during postmoult stages and high during pre-moult. The appendages are replaced rapidly. When they are shed, the concentration of the major moulting hormaone 20-hydroxyecdysone in the haemolymph increases 3-fold.
The exoskeleton of M. rosenbergii contains á-chitin and calcite. Calcite contributes 25% and minor minerals like sodium, potassium and magnesium account for less than 2.5% of total body weight in the prawn. During moulting ionic and osmotic changes take place. In general, crustaceans show a demand for more energy during the process of moulting. This is clear in the case of lobsters, where the glucose level in the haemolymph increases markedly during the pre-moult stage. The subsequent decrease in glucose level or the exhaustion of glucose during post-moult might be due to the utilization of glucose for chitin biosynthesis.
At the time of ecdysis the prawn is inactive and does not feed. It therefore has to utilize reserves, particularly the lipids stored during the prior inter-moult stage. In crustaceans the main storage organ for lipid is generally regarded as the hepatopancreas. This is under the control of the endocrine system. The lipid content will increase at mid pre-moult. A higher level of dietary cholesterol can give an increased moulting rate with a shorter inter-moult stage and might also result in reduced growth. If there is starvation during or after moulting, the larvae can use protein for energy, in case there is lack of stored lipid.
The immune response in the prawns is higher during the inter-moult stage and lower during pre-moult and post-moult stages. The risk of infection is also raised during moulting as the exoskeleton forms a structural and chemical barrier to pathogens and they may gain entry into the body during moulting if there is any damage to the new structures.
Moulting Behaviour
Crustaceans may exhibit extraordinary behaviour during moulting. Shrimps perform intense swimming, walking, jumping, flicking, rolling and rotating movements prior to ecdysis for easy removal of the old exoskeleton. Immediately after the moult the shrimps were found to lie on their side for 5 to 30 minutes. Some Penaeids, for example P. merguiensis and P. esculentus, have been observed to behave similarly.
Change in feeding behaviour
As observed above, prawns do not feed during moulting. A clear understanding of feeding preference and behaviour is important in aquaculture to manage feeding efficiently. This will help minimize food wastages and so protect the pond ecosystem. The feeding response varies, with some activity except during the brief duration of ecdysis itself. Normal feeding is described as an initial stationary searching movements then an approach towards the food target for eventual capture. During ecdysis, the exoskeleton is too soft to permit this activity. Immediately after ecdysis, in most of the cases the prawns eat the exuvia, which is rich in chitin. This will compensate for the loss of minerals due to moulting. Therefore addition of any mineral additives should be applied carefully in the wake of hyper-mineralisation in the water.
Starvation in crustaceans interrupts the moult cycle until food becomes available.
Aggressive behaviour
Social interaction amidst individuals has an effect on growth. In farming ponds, the late pre-moult and early post-moult prawns avoid hard inter-moult and early pre-moult prawns by burrowing into deep, soft mud.
The social interaction leads to the emergence of two groups in the population that can be described as ‘Jumpers’ and ‘Laggards’. Jumpers are the fast growers, with an increased moulting frequency and laggards the slow growers, with different behavioural patterns. This effect, leading to an increased skewness in the size distribution of the prawns, is greater at higher stocking density. Why this should be is not entirely clear. However the competition for limited resources like food and space forms the basis for this phenomenon in M. rosenbergii. Jumpers show an almost 6-fold higher weight increment per moult cycle than non-jumpers. By segregation of the groups farmers can improve performance and increase yield from their stock. The removal of jumpers will trigger the growth in laggards.
Aggressive behaviour has been observed among the prawns during moult cycle, and therefore provision of shelters is greatly recommended in the culture pond. Installation of these artificial habitats in prawn ponds not only increases the surface area for prawns but also increases survival by affording hiding places for moulted prawns, which are prone to be attacked and eaten by hard inter-moult prawns.
Overstocking and paucity of food may also cause cannibalistic behaviour. M. rosenbergii is territorial to some degree and exhibits aggressive and social behaviour. In M. rosenbergii the removal of claws shortened the inter-moult period.
Conclusion
Studies on moulting behaviour are essential for efficient management of prawn farming. In addition, future research should be focused on issues like exploring the causes of ‘Exuvia entrapment disease’ or ‘Moult death syndrome’ in larval stages of prawns, and the use of dietary supplements to synchronise moulting in a particular period to avoid cannibalism, thus mitigating moulting stress to sustain and enhance prawn production.
July 2008