Humans have long exploited the margins of pelagic ecosystems,
but only over the past half century has rapid technological
development allowed fisheries to operate regularly beyond the
sight of land and exploit vast populations of oceanic fishes
that were relatively untouched. Fifty or more years later, the
global impact of fishing on pelagic fishes and their ecosystems is
only now beginning to be understood. Tunas and their relatives,
which include 51 species of tunas, Spanish mackerels, bonitos,
and mackerels (collectively known as scombrids), are major
components of pelagic ecosystems, being both important predators
and forage species that are widely distributed throughout the
temperate and tropical epipelagic waters of the worlds oceans. The majority of tunas and their relatives are highly
migratory with widespread oceanic and coastal distributions;
therefore, their management and conservation are under the jurisdiction
of several international management organizations,
such as the tuna regional fisheries management organizations
(RFMOs). These predators and forage fishes
support some of the largest and most valuable of the worlds
fisheries, sustaining industrial and artisanal fisheries throughout
their ranges, and comprise 12% of global capture fisheries worth
US$ 5 billion a year.
Given the ecological, social, and economic importance of
tunas and their relatives, one might expect that their status and
trajectories would be closely monitored and well understood,
particularly in an era of monitoring progress toward global
biodiversity targets. However, the scale and extent of the global fishing impacts on these important species are surprisingly
uncertain. In 2003, one of the first syntheses brought the
plight of ocean predators (mainly tuna species) to the attention
of the wider scientific community, concluding that global community
biomass of large pelagic fishes had been reduced by
around 90% from preindustrial abundance. However, this
work relied heavily on an analysis of catch and effort data from
only one fishing gear type, resulting in an overestimation of tuna
declines. Alternatively, fisheries stock assessments provide
a more reliable estimate of population size and trajectory
and are regarded as the preferred source of information with
which to assess the effects of fishing on fish populations and
ecosystems. In light of the problems with catch data, the
increasing availability of stock assessments, and increasing public
concern for the sustainable long-term management of natural
resources, it seems timely to evaluate the global development,
trajectory, and sustainability of tuna fisheries and their relatives.
Here, we evaluate the trajectory and exploitation status of 26
populations of tunas and their relatives. First, we quantify the
overall impact of fishing on adult biomass globally, including the
extent of the impact within major oceans, major taxonomic
groups, and species with different life history strategies using two
metrics: the average annual rate of change and the total extent
of decline. Second, we compare the adult biomass trajectories
against the current exploitation status of each population determined
by two standard biological reference points: the ratio
of the current adult biomass relative to the adult biomass that
would provide the maximum sustainable yield (MSY; B/BMSY)
and the ratio of current fishing mortality relative to the fishing
mortality rate that maintains MSY (F/FMSY).
Results
We assembled age-structured stock assessments with >15 y of data for 17 tuna populations (7 species), 5 mackerel populations (2 species), and 4 Spanish mackerel populations (2 species) of the 51 species of scombrids. We observe that those mackerel and tuna species with the largest number of stock assessments are the most economically important species, comprising 70% of the global reported catches (Fig. 2A). In contrast, the small tunnies, Spanish mackerels, and bonitos, which are mainly tropical coastal species, have a smaller number of stock assessments available. The status of these tropical coastal scombrids is mostly unknown throughout their ranges, despite the importance of their commercial fisheries for many coastal fishing communities in many developed and developing countries around the world.
Trajectories of Catches and Adult Biomass Across Tunas and Their Relatives
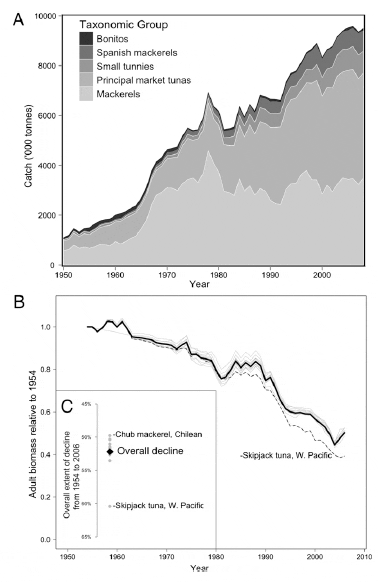
The annual catches of tuna and their relatives have
risen continuously since the 1950s, reaching 9.5 million tonnes in
2008 (Fig. 2A). This increase in catches was achieved by halving
global tuna biomass in half a century; total adult biomass summed
across all monitored populations has declined globally by 52.2%
from 1954 to 2006 (Fig. 2 B and C). This total extent of decline
depends on the inclusion of the most abundant populations, and
their contribution to the overall decline can be seen by excluding
a single population at a time from the analysis and recalculating
the overall trend (jackknifing). The overall extent of decline
would have been 8.2% greater (60.4%) were it not for the inclusion
of the abundant West Pacific skipjack tuna population.
The overall annual rate of decline can be calculated from the
yearly (i) changes in biomass (ri,j) among populations (j), which
accounts for temporal autocorrelation and the wide variation in
the absolute size of populations (Methods). On average, the annual
rate of change in adult biomass was -1.7% y-1 [95% confidence
interval (CI): -2.6 to -0.9] across the 26 populations
from 1954 to 2006. This global annual rate of change is equivalent
to an average decline of 59.9% across all populations within the 52-y period. Moreover, the trajectories in adult biomass of tunas
and their relatives vary widely across oceans, taxonomic groups,
species, and life history strategies.
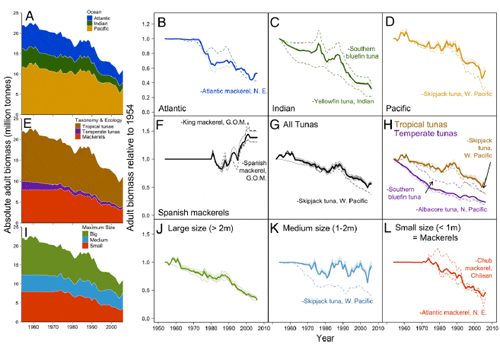
The total extent of decline in adult biomass has been greatest
in the Indian Ocean, with a 63.6% decline from 1954 to 2006,
compared with a 49.6% decline and a 49.2% decline in the Atlantic
and Pacific Oceans, respectively (Fig. 3 AD). In the Pacific
Ocean, the catches of tunas and their relatives are dominated by
the abundant West Pacific skipjack tuna adult biomass, which
comprises 64% of the total tuna catches in the western Pacific
Ocean. After excluding West Pacific skipjack, the extent of decline
in adult biomass in the Pacific Ocean is 66.6%. Therefore,
the large observed declines in adult biomass suggest substantial
impacts of fisheries in all three oceans, despite the different
timing in the historical expansion of industrial fisheries. Industrial
fisheries, particularly those targeting tuna species, started in
the 1950s and 1960s in the Atlantic and Pacific Oceans, whereas
they fully developed two decades later in the Indian Ocean. We
also observed that the fastest annual rates of decline within the
52-y period occurred in the Indian Ocean (-3.2% y-1, 95% CI:
-4.8 to -1.6), possibly attributable to
aggressive and poorly regulated artisanal and industrial fisheries
operating in a relatively ligh tly exploited ocean.
Of the three major taxonomic groups of tunas and their relatives,
only the total adult biomass of all Spanish mackerels has
recovered, increasing by 38.2% over the past half century (Fig.
3F). The status of the four Spanish mackerel populations off
the southeastern coast of the United States is currently healthy
following the implementation of a recovery program after many
years of overfishing. Of the other two taxonomic groups, the
total adult biomass of all mackerels has declined the most
(58.1%), whereas tunas have declined by 49.1% (Fig. 3 G and L).
However, after excluding the abundant West Pacific skipjack
tuna, the total biomass of all tunas has decreased by 62.5% from
1954 to 2006.
The life history and ecology of fishes are intimately linked to
their response to exploitation. Larger species tend to be preferentially
targeted by fisheries over smaller species and may be
intrinsically more sensitive to fishing because of their relatively
less productive life histories. However, this ecological pattern
can be overwritten by aggressive globalized fisheries.
We observed that the total adult biomass of the largest species,
bluefins, bigeye, and yellowfin tunas, and the smallest species,
mackerels, has declined the most, 62.8% and 58.1%, respectively,
since 1954 (Fig. 3 IL). In addition, we only found significant
and steep rates of decline in adult biomass in the largest
species, -2.4% y-1 (95% CI: -3.5 to -1.4).
We hypothesize that the large interannual variability observed
in the adult biomass trends of the smallest pelagic coastal species
may be hindering the detection of significant declines in their
overall annual rates of change.
We also find that the biogeography of tuna life histories matters.
Temperate tuna populations have declined more steeply,
-3.1% y-1 (95% CI: -4.2 to -1.9), than tropical tuna populations,
-1.7% y-1 (95% CI: -2.8 to -0.7). These rates are equivalent to an average decline of
80.2% across all the temperate tuna populations and 59.5%
across all the tropical tuna populations. Temperate and tropical
tuna species have biogeographically distinct life history strategies:
temperate species (bluefins and albacore tunas) are longer lived,
reproduce later, and have a shorter breeding season and a geographically
more restricted breeding site than the tropical tuna
species (yellowfin, skipjack, and, to some extent, bigeye tunas),
making them more accessible to fisheries, and therefore overall
less productive fisheries.
Link Between the Adult Biomass Trajectories and the Current Exploitation Status
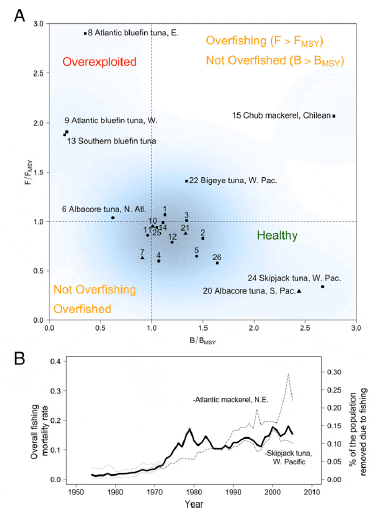
Population and species trajectories depend not only on life histories and ecology but on the level of exploitation. Here, we summarize the current exploitation status for the 21 populations for which we were able to obtain estimates of the two biological reference points, B/BMSY and F/FMSY (Fig. 4A). We define overfished to mean that the biomass of the population has been reduced to a level less than that which would provide the MSY (B < BMSY) and the term overfishing to mean that a population is being subjected to a fishing effort greater than that required to produce the MSY (F > FMSY), a definition used by the majority of the tuna RFMOs. First, there are a total of 4 overexploited temperate tuna populations that are overfished and are experiencing overfishing: East and West Atlantic bluefin tunas, Southern bluefin tuna, and North Atlantic albacore tuna (Fig. 4A). Second, there are 12 populations, mostly tropical tunas and Spanish mackerels, currently considered healthy (B > BMSY and F < FMSY). Finally, there are 5 populations of tunas and mackerels in an intermediate state that either have a biomass below healthy levels or a fishing mortality exceeding healthy levels but not both (B < BMSY or F > FMSY). Although the current exploitation status of tunas and their relatives can be easily categorized according to their biological reference points, it is important to highlight that the majority of tunas and their relatives, despite their assigned exploitation status, have been fished down to around MSY levels, and are therefore fully exploited. The extent of the declines in adult biomass is consistent with the current exploitation status of the populations; the populations having experienced the largest declines in biomass are either fully exploited or overexploited.
Discussion
The global adult biomass of tunas and their relatives has been
halved over the past half century but not without yielding considerable
catches, income, and food for the benefit of humanity.
However, these population declines cannot continue without
compromising yields in the near future: The majority of populations
are fully exploited, which limits the further expansion of
catches from these fisheries. Currently, fisheries catch around
1015% of the tunas and their relatives each year globally. The global demand for tunas and
their relatives is still increasing, as is the trajectory of fishing
mortality.
The largest declines in adult biomass have occurred in two
groups of species with distinct life histories, the largest and less
productive temperate tunas and the smallest and more productive
mackerel species. Mackerels would, a priori, be considered
intrinsically resilient to overfishing because of their fast
life histories, being fast-growing, early-maturing, and short-lived;
yet, mackerels exhibit some of the steepest declines. However, it
has been shown that within the past 50 y of industrial fisheries,
the collapse of small-and fast-growing pelagic species has been
more frequent than in larger species . Since fisheries developed
in the 1950s, they have preferentially targeted largebiomass,
shallow-water species, such as small pelagics. This
historical pattern of fisheries development, combined with the
increasing global market demand for small pelagic fish as food,
fishing bait, fish meal, and oil, has probably contributed to
their massive declines. The role of life histories is more apparent
in tunas. The less productive temperate tuna species have been
affected the most by fishing, exhibiting steeper and larger declines
than the more productive tropical tuna species, suggesting
that low productivity and slower life histories might be an important
factor, together with catchability, accessibility, and market
price and demand, in determining the vulnerability of the
species to fishing.
The reductions in adult biomass of tuna populations estimated
in our global analysis differ from the more pessimistic interpretations
of the global status of tuna fisheries described by Myers
and Worm. Although the two studies are not strictly comparable,
Myers and Worm found a 90% decline, on average, in
the catch per unit effort of large pelagic fish species and we found
a 59.9% decline, on average, in the adult biomass of tunas and
their relatives. Notwithstanding the gross differences, both studies
agree on the steep declines of three bluefin populations and
one albacore population, which are clearly overfished with current
biomasses below BMSY. Instead, our results present a wide
range of trajectories across tuna populations, which are more
consistent with the findings of a study by Sibert et al., which
reports declines ranging from 1188% from baseline adult biomass
across the Pacific tuna populations. Moreover, our findings
are consistent with those of a recent evaluation of the global
conservation status of scombrid species carried out by the International
Union for Conservation of Nature (IUCN), which
showed that 68% (35 of 61 species) of scombrids are not considered
to be threatened with extinction but that a few (5 species)
have declined sufficiently to trigger listing under the IUCN Red
List Threatened categories, notably the Southern and Atlantic
bluefin tunas. We caution that our estimates of total and
average declines in adult biomass are almost certainly an underestimate,
because fishing began long before the start of many
of the time series summarized here. Stock assessments often
begin years after the start of a fishery and may even be triggered by declining catches, as, for example, in the case of the Atlantic
bluefin tuna, which was essentially fished out in the South Atlantic
in the 1960s before formal assessment. Finally, we also
show that, globally, the majority of the tunas have been already
fished down to near MSY-related levels. From a fisheries
management perspective, MSY is usually obtained when the
biomass of a population has been reduced by 6070%.
Nonetheless, from a conservation perspective, the 52.2% global
decline in total adult biomass and the average population declines
of 59.9% across tunas and their relatives increase the probability
of ecological and economic extinctions of target populations, with
considerable biodiversity consequences for bycatch species.
In addition, the magnitude of these declines creates concerns
about the potential unknown ecosystem effects of removing large
amounts of biomass from the pelagic food webs.
MSY is the explicit or de facto target yield level for most tuna
RFMOs. Given that 4 of the 26 populations are substantially
below BMSY (Fig. 4A) and the others are all at target levels larger
than 0.9BMSY, most fisheries managers would consider these to
be extremely well managed (with the exception of the 3 bluefin
and 1 albacore populations). However, there is little room for
complacency. We highlight three issues to be tackled with urgency
to reduce the risk of tropical tunas and other scombrid
populations deteriorating in the same way as the bluefin tunas
and to minimize the considerable collateral damage and biodiversity
consequences of these fisheries. First, tuna productivity
is apparently declining; the current estimates of MSY for some
tuna populations are lower than in the past, partly a result of the
increased mortality of immature tunas in the past two decades
from purse seine fisheries, which has consequently decreased the
maximum potential yield of the fisheries. Second, their high value and global demand, and the rising fishing capacity and
mortality (Fig. 4B), are exacerbating the pressure on
populations that are already fully exploited or, in some cases,
overexploited. Management of tuna populations under the single-
species approach appears to be largely successful for the less
valuable tropical species but has not been effective for high-value
bluefin tunas driven by the scale of international demand for and
trade of high-valued tunas. In those cases, additional measures
seem to be required. Here, we have a case where trade is overwhelming
the, normally effective, scale of fisheries management.
Hence, there appears to be a role for conservation tools, such
as the Convention on International Trade in Endangered
Species (CITES), to work alongside the existing management
framework to ensure the recovery and future sustainable fishing
of the most exploited populations. Third, exploitation of productive
species, such as tunas, at MSY is driving steep population
declines and elevating the risk of extinction of some unmanaged
and less productive bycatch species. Tuna fisheries are directly
responsible for endangering a wide range of oceanic pelagic
sharks, billfishes, seabirds, and turtles.
Many of these issues could be alleviated if fisheries management
organizations treated MSY as an upper limit rather than a
target reference point in their management objectives, a longstanding
recommendation of several international United Nations
Food and Agricultural Organization agreements and guidelines
over the past 15 y. Most tuna RFMOs have vague management
objectives and have not adopted or implemented specific
targets and limits. We recommend the development of
well-defined management strategies involving harvest control
rules and the associated decision rules that can keep the fishery
within defined limits. These would potentially facilitate the creation of well-defined and specific targets and limits for each population
(and therefore management objectives), improving the
decision-making process and speeding the implementation of
appropriate management measures. The use of upper
limits and lower targets would improve profitability and reduce
the impacts on ocean biodiversity.
The long-term sustainability of tunas and their relatives can only come from stricter management measures to treat MSYrelated levels as a limit rather than a target management objective, to reduce the overall fishing capacity, and to rebuild overexploited populations, as well as further implementing regulations to minimize the collateral impacts of these fisheries on marine ecosystems.
Further ReadingYou can view the full report and list of authors by clicking here. |
January 2013


