Due to these severe negative effects, a number of strategies have been proposed to control HABs, involving ultraviolet light, microwave, clay, modified sand, plants, protozoan, and Chinese traditional medicines. However, only a few of these are feasible and applicable in case of emergencies due to the high cost or side effects. In recent years, microbial agents mitigating HABs, especially naturally occurring algicidal bacteria have attracted global attention.
Algicidal bacteria play a potentially important role in regulating the growth, metabolism, and toxin production of harmful algae. Factually, relationships between algicidal bacteria and harmful algae are quite complex and have been extensively researched, of which the most conspicuous and important is the inhibition or lysis of harmful algae by algicidal bacteria. Consequently, plenty of algicidal bacteria which mainly belong to genera Pseudoalteromonas, Alteromonas, Vibrio, Cytophaga, and Saprospira were isolated. The negative effects of algicidal bacteria on harmful algae are therefore considered as the basic principle of microbial methods to control HABs. The mode of action of algicidal bacteria can be divided into direct attack and algicide release. Direct attack requires attachment of bacteria to the harmful algae in order to make direct lysis, while algicide release mode is that bacteria release freely diffusible algicides, such as protein, amino acid, or alkaloid, to kill algal cells. With regard to the relationships, another aspect is that some harmful algae may inhibit or lyse algicidal bacteria, while some may supply nutrition to algicidal bacteria. What is more special is that the co-culturing of algae with bacteria can enhance the production of algicidal compounds derived from algae themselves. Besides these interactions above, bacteriophages are supposed to be involved in the relationships, through lysing algicidal bacteria to trigger the growth of harmful algae. Presently, the major targeted harmful algae that have been investigated are dinoflagellates, diatoms, and cyanobacteria.
Phaeocystis globosa, a eukaryotic HABs-causing species, is reported to be associated with shellfish mortality, acid rain, and serious impacts on the whole ecological environment by releasing toxic hemolytic substances, forming nuisance foam, and causing oxygen depletion. Meanwhile, the extracellular polymeric substances of P. globosa were also involved in the stability of quantum dots (toxic nanoparticles) in marine environments. P. globosa blooms have frequently occurred along the southern coast of China, the eastern English Channel, and the South Central coast of Viet Nam. However, only a few studies have focused on the interactions between P. globosa and microbes (bacteria and viruses) in recent years. Even though actinomycetes are commonly known for production of bioactive compounds, a few of algicidal actinomycetes and their algicidal activity against P. globosa were reported in attempt to develop potential microbial control methods.
In this study, we isolated an actinomycete strain RPS algicidal to P. globosa from Fujian Zhangjiangkou Mangrove National Nature Reserve in China. This actinomycete was identified as Streptomyces alboflavus. Furthermore, the algicidal mode, sensitivity of algicidal substance to temperature and pH, molecular weight range and polarity of the active substance, algicidal process of the ethyl acetate extract, and susceptible algae range were investigated to better understand the novel activity against P. globosa.
Results
Isolation of algicidal actinomycete RPS
The strain named RPS was isolated from the sediment of Fujian Zhangjiangkou Mangrove National Nature Reserve. Algicidal activity of RPS was tested in a 24-well cell plate (Figure 1). This figure indicated that the biomass of the algal culture treated with 150 μL, 200 μL, and 300 μL of RPS decreased dramatically in 48 h, while the control grew normally. A significant interaction effect of RPS supernatant and time (P<0.001) was also observed by analysis of two-way ANOVA. Nearly all algal cells fell to the bottoms of the wells after 4 h. Combined with cells sinking, there was approximately an 100 per cent algicidal ratio at 48 h which proves that strain RPS is a strong algicidal actinomycete against P. globosa. Since treatments with 200 μL and 300 μL of supernatant showed almost the same activity during 48 h (P>0.05), 200 μL was chosen as the basic amount to determine the culture activity in different treatments in the following studies.
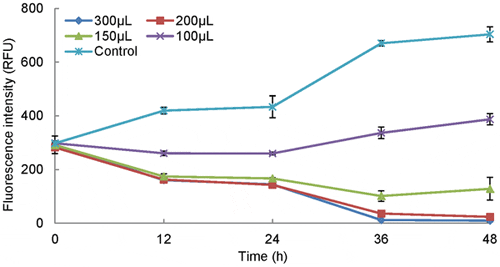
Data points: mean ± standard deviation of triplicate assays. Error bars not visible are smaller than symbols. The algal biomass treated with RPS supernatant decreased dramatically in 48 h. A significant interactive effect of RPS supernatant and time was also observed by analysis of two-way ANOVA (P<0.001).
Identification of algicidal actionmycete RPS
The colonies of strain RPS on Gause synthetic agar No. 1 medium for 15 d were circular and dry. The substrate mycelia were abundant and golden-yellow, while aerial mycelia were rare. No soluble pigments were released into the medium. Spores were cylinder with a smooth surface (Figure 2A). Typically, all these characteristics belong to Streptomyces. Cultural characteristics of aerial mycelia, substrate mycelia, and soluble pigments on usual media were similar with each other (Table 1). Substrate mycelia were abundant and yellow or red in color, while the aerials were poor and in pale color (white or light pink). No pigments were released by RPS growing on all media except potato dextrose agar (light yellow).
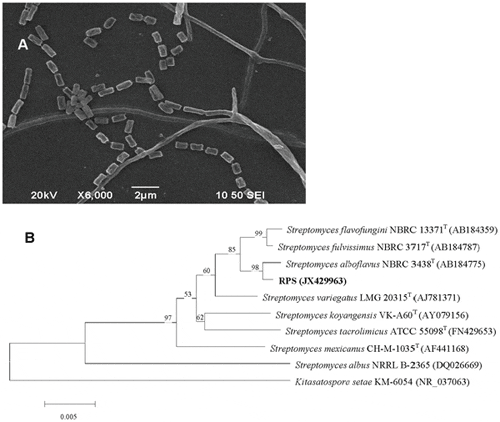
Morphology from SEM results (JSM-6390, JEOL) and phylogenic tree based on 16S rDNA sequences by Neighbor-Joining analysis. Kitasatospora setae KM-6054 (NR_037063) was used as the out group.
Table 1. Culture characteristics of RPS at 28°C on different media.

In terms of physiological and biochemical characteristics, RPS was capable of liquefaction of gelatin, peptonization of milk, and hydrolysis of starch, and able to use glucose, sucrose, glycerol, galactose, mannitol, and xylose as the sole carbon and energy source, but could not utilize cellulose, lactose, or maltose to grow.
The 16S rRNA gene of RPS was amplified, and an approximately 1.5 kb band was observed by 1 per cent agarose electrophoresis. The obtained sequence (GenBank accession number JX429963) showed that the most probable affiliation of RPS was to the genus Streptomyces by BLAST, and the strains with similarity of more than 98 per cent were all streptomycetes. The phylogenetic tree (Figure 2B) produced by the neighbor-joining analysis revealed that the closest relative of RPS was the strain S. alboflavus NBRC 3438T with the similarity of 99.7 per cent, and all streptomycetes were classified into the same branch.
RPS was very similar to S. alboflavus in morphological, physiological, and biochemical characteristics, including poor aerial mycelia, yellow to red substrate mycelia, no soluble pigments released, potential of liquefaction of gelatin, peptonization of milk, and hydrolysis of starch [42]. In further consideration of the close phylogenetic relationship to S. alboflavus, strain RPS was identified as Streptomyces alboflavus (S. alboflavus RPS).
Algicidal mode
Algicidal activities of initial fermentation broth, supernatant, and mycelia were investigated. The algicidal ratios of the initial broth and supernatant reached 99 per cent and 95 per cent (a same active level P>0.05), respectively, while mycelia showed a much lower ratio of approximately 36 per cent (P<0.001, Figure 3). The significant difference (P<0.001) of the algicidal activity from this comparison shows that RPS exhibits the algicidal activity in an indirect way by releasing an extracellular active substance into the growth medium. The partial activity in the mycelia treatment may be caused by some active substance released by mycelia after being added into the algal culture.
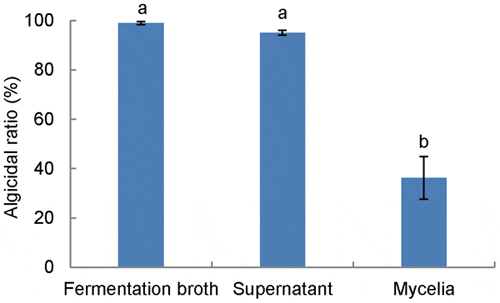
Data from RFU assay of algicidal effect (1:10, v:v). Data points: mean ± standard deviation of triplicate assays. Treatment means with different letters differed significantly (P<0.001), analyzed using one-way ANOVA.
Sensitivity of algicidal activity to temperature and pH
The supernatant exhibited different trends in treatments with different temperatures and pH for 2 h. The activity of the supernatant was very sensitive to temperature, as it declined significantly (p<0.001) by 25 per cent and 80 per cent when the temperature was raised to 50°C and 60°C (and above), respectively, compared with the untreated supernatant (Figure 4A). However, the activity was resistant to pH, as the supernatant was similar active (p>0.05) in a pH range of 3–11 (Figure 4B). Hence, to keep the activity in the supernatant, more attention should be paid to the steps related to temperature changes.
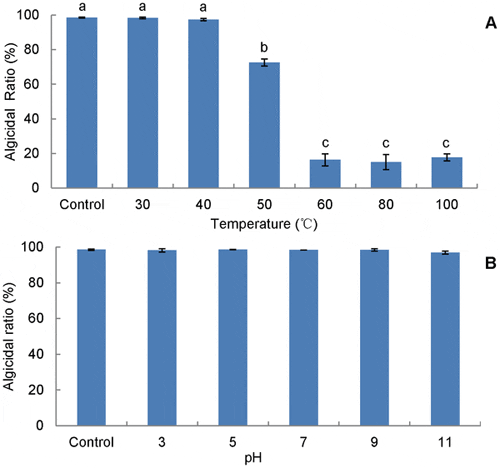
Data from RFU assay of algicidal effect (1:10, v:v). Control represents the initial fermentation broth without any treatments. Data points: mean ± standard deviation of triplicate assays. Temperature treatment means with different letters in (A) differed significantly (P<0.001) by analysis of one-way ANOVA, suggesting that the algicidal activity of RPS is sensitive to temperature, while no significant difference (P>0.05) was observed among treatments with pH in (B), meaning that the activity is insensitive to pH.
Molecular weight range and polarity of algicidal substance
The algicidal activities of dialyzed supernatants were significantly different (p<0.001) from each other (Figure 5A). The figure showed similar activities (p>0.05) in the initial (91 per cent) and dialyzed supernatant with a cut-off of 100 Da (89 per cent), but dramatically decreased activity (p<0.001) in the dialyzed supernatant with a cut-off of 1000 Da (10 per cent). Certain reduction (p<0.001) was also observed in dialyzed supernatant with a cut-off of 500 Da (not as dramatic as that of 1000 Da). Therefore, it is credible to speculate that the molecular weight of the active substance ranges between 100 Da and 1000 Da.
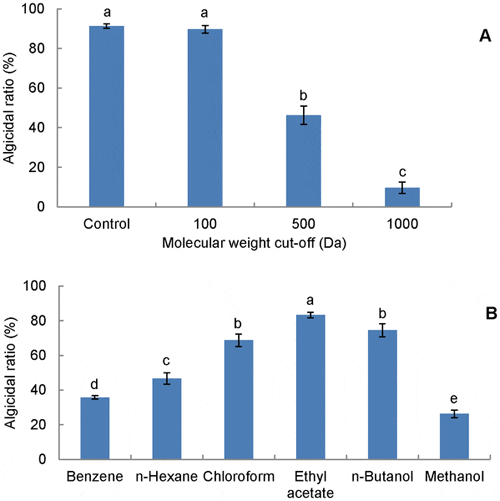
Data from RFU assay. Data points: mean ± standard deviation of triplicate assays. In (A), initial supernatant (Control) and dialyzed supernatants were added at 10 per cent ratio, and treatment means with different letters differed significantly (P<0.001) by analysis of one-way ANOVA. In (B), 10 mu;L extract was added into 1.99 mL algal culture (equal to 1:10 of v:v in algicidal substance dose, no loss considered during extraction), and treatment means with different letters differed significantly (P<0.05) by analysis of one-way ANOVA.
The polarity of the active substance was investigated through the comparison of activities of different organic extracts (Figure 5B), of which the ethyl acetate extract possessed the highest activity (83 per cent). Other solvents, which were more or less polar than ethyl acetate, led to lower extraction efficiencies (p<0.05). These results imply that the active substance should be a low polar and hydrophobic molecule with a low molecular weight. As the extract amount (10 μL) added in this experiment was equal to 200 μL of initial culture with no loss considered during extraction, ethyl acetate was able to extract approximately 90 per cent of the active substance in the supernatant.
Algicidal process
Nearly all algal cells fell to the bottoms after 4 h treatment with ethyl acetate extract, and the precipitated algal cells faded gradually along with time. The whole algal culture was clear almost without color at 48 h. Based on the SEM observation and optical observation of several time-point samples, the algicidal process was inferred as follows. A normal P. globosa cell possesses two intact flagella (Figure 6A); the movement of alga was inhibited by the active component added, which even caused the flagella to fall off the algae (Figure 6B); the active component ruptured the algal cells, leaving broken cell walls (Figure 6C); as not all cells were sensitive to the active substance at the same level, the former three occasions might exist in the same field of vision (Figure 6D). Factually, cell lysis was also proved by the results in Figure 1, where the fluorescence intensity dropped dramatically over time.

(A) normal P. globosa, (B) flagella falling off the alga, (C) broken cells, and (D) algae at different damaged levels. The putative algicidal process is that the algicidal substance inhibits the movement of P. globosa, then causes the flagella to fall off the algae, and finally lyses the algal cells.
Susceptibility of algae to RPS
The ethyl acetate extract lysed 10 of the 14 tested algae (Table 2), which were raphidophyte P. globosa, yellow-green algae of H. akashiwo, green algae of Chlorophyta (P. helgolandica, P. subcordiformis), golden algae of Chrysophyta (D. inornata, I. galbana), dinoflagellates of Pyrrophyta (A. tamarense, S. trochoidea), and diatoms of Bacillariophyta (P. tricornutum, A. japonica). However, green algae of Chlorophyta (C. autotrophica, N. oceanica, D. salina, Chlorella sp.) were not lysed. These results indicate that RPS has a wide target range and certain specificity, since it had no effect on 4 tested green algae (Table 2).

Discussion
Algicidal Streptomyces against harmful P. globosa
Phaeocystis blooms have occurred frequently around the world since its first recorded observation as early as 1923. These blooms exerted negative effects on higher trophic levels in the marine ecosystem, and consequently influenced human activities such as fisheries and coastal tourism. Hence, it is urgent to investigate the interactions between microbes and Phaeocystis, and develop potential microbial control methods. In this study, we isolated and identified Streptomyces alboflavus strain RPS, and then focused on its strong algicidal function and novel algicidal properties against P. globosa. Indeed, several strains of actinomycetes have been reported to show antialgal activity against cyanobacteria and dinoflagellates. However, there have been a relatively few algicidal Streptomyces strains reported against P. globosa, including the strain of Streptomyces O4–6 as well as two bacteria belonging to another genus Bacillus. Therefore, the study of RPS will further widen the resource pool of antagonistic microbes to P. globosa, as well as other harmful algae like A. tamarense and H. akashiwo (Table 2).
High algicidal activity of RPS
S. alboflavus RPS achieved a 95 per cent algicidal ratio against P. globosa in a short time of 48 h (Figure 1), which proves RPS is a highly efficient algicidal streptomycete. Previous reports showed much weaker algicidal effects. For example, Streptomyces S-9 isolated by Yamamoto et al. needed 5 d or more to achieve a stable effect on cyanobacteria. Also, Choi et al. reported that the cyanobacterial biomass was just suppressed by 84.5 per cent after 7 d treatment of S. neyagawaensis, and Yan et al. showed that bacteria Y01 and Y04 could remove P. globosa in 6 d. In this study, the similar activity (p<0.001) in the initial broth and supernatant after centrifugation indicated that RPS exerted the algicidal activity through the algicide release mode (Figure 3). Although algicidal compounds have been investigated for decades, leading to identification of algicidal proteins [20], peptides, amino acids, antibiotics, pigments, alkaloids, and biosurfactants, difficulties still exist in the isolation and elucidation processes of the algicidal compound in this study (We got a fraction with 99 per cent algicidal ratio at 20 μg mL−1 through silica column chromatography, TLC, and HPLC). Algicidal compounds and their bioactive characteristics vary greatly between species of algicidal bacteria, requiring different and specific compound isolation procedures both in extraction and chromatography.
Novel algicidal features
RPS showed several novel algicidal features. What should be first focused on is the sensitivity to temperature and pH (Figure 4). Treatments at 50°C and above resulted in vast loss (p<0.001) of activity of the supernatant (Figure 4A), which led us to think the active component could be a protein, as several previous algicidal proteins were reported to be susceptible to temperature. For example, Lee et al. reported an extracellular protease from Pseudoalteromonas A28 against Skeletonema costatum and the protease activity was abolished by incubation at 68°C for 1 h or 100°C for 15 min. Also, Paul and Pohnert reported an algicidal protease lost the activity against S. costatum in the condition of 80°C for 10 min. However, the stable activity (p>0.05) of RPS within the pH range of 3–11 (Figure 4B) implies that the active component may not be a protein, as active proteins are generally pH-sensitive, such as violaxanthin deepoxidase, potassium channel, and Glycoprotein G. What's more is that the treatment with proteinase K did not reduce the activity of RPS supernantant (data not shown). This paradoxical evidence suggests that this strain possesses a unique active substance which is temperature-sensitive but not pH-sensitive.
The dialysis result revealed that the molecular weight of the active substance should range between 100 Da and 1000 Da (Figure 5A), meaning a small molecule. Recently, several small molecule algicides have been reported, including β-cyanoalanine, nigericin, harmane, and rhamnolipid biosurfactants. As for the partially reduced activity (p<0.001) in dialysis bag of MWCO of 500 Da, it is possible that the molecular weight of the active component is close to 500 Da, meaning that some of the component could occasionally pass through the dialysis bag with the cut-off of 500 Da and cause the decrease in activity. Extraction by ethyl acetate recovered nearly 90 per cent of active substance in the supernatant, and showed the highest activity (p<0.05, Figure 5B). This high efficiency indicates that ethyl acetate can be used to extract the active substance in late-stage study. In fact, organic solvents were often used to isolate algicidal compounds. For example, Sakata et al. isolated the 2,3-indolinedione against Chaetoceros ceratosporum by ethyl acetate. Li and Hu also reported an algicidal ethyl 2-methylacetoacetate separated by ethanol and chloroform.
Based on the sensitivity to temperature, insensitivity to pH, molecular weight range, and high extraction ratio by ethyl acetate, it is inferred that the active component should be a low polar substance with a low molecular weight. To avoid degradation or loss of the bioactive component, more attention should be focused on temperature during the subsequent isolation and conservation steps, including concentration of supernatants, evaporation of extracts and chromatography.
Presumption of algicidal process
Through the SEM and optical observation, it is estimated that flagella movement of P. globosa was efficiently inhibited by the active substance in the ethyl acetate extract, resulting in decreased cell motility. Subsequently, the flagella fell off the cells, and these attacked cells were lysed (Figure 6). This presumption was confirmed by the fact that the algae were sunk in the first 4 h after the addition of the active substance under optical observation, and the amounts of flagella and broken cell walls were observed later under SEM. Additionally, most of the susceptible algae possess flagella, such as yellow-green algae H akashiwo, green algae P. helgolandica and P. subcordiformis, dinoflagellates A. tamarense and S. trochoidea, and golden algae I. galbana (Table 2), while the non-susceptible algae do not possess flagella, including C. autotrophica, Chlorella sp., and N. oceanica (Table 2), which may be an indirect proof that the active component targets the flagella. Although this presumption needs more supporting evidence to prove, it may be a novel direction to study the algicidal mechanisms focusing on the movement of flagella. This direction may hopefully break through the bottlenecks of the present algicidal mechanism studies that have been primarily focusing on the oxidative damage and antioxidant responses, such as CAT, MDA, SOD.
Susceptible algae
RPS showed a wide algicidal range toward harmful algae. RPS was antagonistic to HABs-causing algae P. globosa, H. akashiwo, A. tamarense, S. trochoidea, and A. japonica, as well as some other algae P. helgolandica, I. galbana, etc. However, it was not active against C. autotrophica (fish bait alga), D. salina (fish bait alga), N. oceanica (potential biofuel alga), and Chlorella sp. (Table 2), which may be an advantage (no effect on beneficial algae) of RPS when used as a biological agent in control of HABs in the future. Besides, S. alboflavus RPS possesses the great potential to produce industrial pigments when the culture supernatant is used as an algicidal agent, since there was a large amount of red matter that was separated from the ethanol acetate extract of RPS mycelia.
Conclusion
A strong algicidal streptomycte RPS against harmful algae P. globosa was isolated and identified as Streptomyces alboflavus. Furthermore, the novel algicidal features of RPS were investigated and concluded as follows: (i) high algicidal efficiency of more than 95 per cent algicidal ratio in 48 h; (ii) sensitivity to temperature at and above 50°C, but resistance to a pH range of 3–11; (iii) the molecular weight of active component ranging 100–1000 Da and approximately 90 per cent extraction ratio by ethyl acetate; (iv) putatively algicidal process of inhibiting the movement of flagella, followed by lysis; (v) a wide target range against harmful algae P. globosa, H. akashiwo, A. tamarense, S. trochoidea, and A. japonica. RPS with the high and novel algicidal activity gives a hint on controlling P. globosa blooms by microbial methods and effectively extends the resource pool of algicidal microbes against harmful algae.
May 2014



