Introduction
Methane (CH4) is the terminal product of anaerobic respiration when all electron acceptors except carbon dioxide (NO3δ, Mn+4, Fe+3 and SO4δ2) have been depleted by the microbial community. The production of methane is performed by methanogens, which are microorganisms belonging to the domain Archaea.
In 1906, Sohngen showed for the first time that CH4 can serve as an energy and carbon source for bacteria. Biological oxidation of CH4 is now known to occur aerobically in both terrestrial and aquatic habitats, and anaerobically in sediments and anoxic salt water. The aerobic methane oxidizing bacteria (MOB) have been classified into the phylum Proteobacteria and recently also Verrucomicrobia (based on three strains), the latter representing thermophilic acidophiles and still being under taxonomic debate. The MOB in Proteobacteria have 16 recognized genera within the classes Gammaproteobacteria (traditionally referred to as Type I MOB) and Alphaproteobacteria (Type II MOB), although some genera do not fit these generalizations very well.
Aquatic food webs are supported by organic matter derived from phytoplankton, macrophytes or imported from the surrounding land (allochthonous sources). CH4 production accounts for a large proportion of the total organic matter degradation in freshwater lakes (2056%), and is a source of energy and carbon for aerobic MOB in the water column and the sediment, linking the anoxic and oxic communities.
Isotopic distributions in animals are generally closely related to dietary isotopic composition. The stable isotopic compositions of animal tissues reflect both long-term and short-term diets in slow and fast turnover tissues. Measurement of carbon stable isotope ratios (δ13C) has been a successful tool in food web studies. Because CH4 is much depleted in 13C, low δ13C values in organisms of aquatic food webs have been considered to indicate their consumption of CH4-derived carbon. Such low values of δ13C in animal biomass have been reported for invertebrates in various aquatic systems.
Low δ13C values have been reported for fish, and a few studies have suggested that CH4-derived carbon could be transferred in substantial amounts to higher trophic levels. Calheiros (unpublished data) found strongly negative δ13C values for zooplankton, aquatic insects and detritivorous fishes in a Brazilian Pantanal lake, being the first study to discuss the importance of methanotrophs for the whole food web in this wetland.
While more negative δ13C values can be an indicator of CH4 carbon, the isotopic signal from other potential sources of 13C-depleted carbon such as primary production based on CO2 from respiration can provide alternative explanations. Therefore, the combination of stable isotope analysis with other independent biomarkers is needed to elucidate if low δ13C values are due to assimilation of carbon from biogenic CH4.
The lipids of the proteobacterial MOB have special fatty acid (FA) composition. They contain a few very unusual FAs that have been used as efficient group-specific markers in studies of abundance and dynamics in methanotrophic community structure. A fraction of the assimilated FAs of prey organisms is stored in cells rather than being degraded and the chain lengths and double bond positions in such stored FAs are preserved. On a general level, variation in the FA composition using specific MOB FA can be used as a traceability tool for determining the relative dependency on bacteria versus phytoplankton in diets [34] and seasonal variation in food quality used by zooplankton.
Similarly, monounsaturated fatty acids, like those diagnostic for MOB, can thus be microbial biomarkers indicating transfer of carbon and energy from methane to higher food web levels. The assumption that animals actually ingest MOB has been supported by the detection of phospholipid fatty acids (PLFAs) diagnostic for MOB in tissues of chironomid larvae. In addition, Deines et al. experimentally confirmed that MOB carbon can be transferred to invertebrate animals. In this way, findings that FAs specific for MOB are present in tissues of 13C-depleted freshwater invertebrates have been supporting the hypothesis of a link between MOB and animal food webs. However, MOB specific FAs have not been reported in fishes so far.
In this study, we tested the hypothesis that methane carbon from MOB can be transferred through food webs all the way to fish, by combining analyses of δ13C and fatty acid composition in fish, benthic macroinvertebrates, zooplankton, and the dominating aquatic macrophyte from a shallow tropical lake in Pantanal, Brazil. We found specific fatty acids of methane-oxidizing bacteria in tissues of aquatic invertebrates and fish, showing that methane can in fact contribute carbon to large parts of aquatic food webs and production of fish biomass.
Results and Discussion
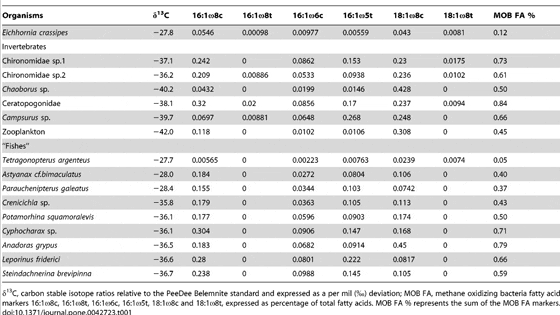
All δ13C values ranged from -42.0 to -27.7 and the contribution of MOB FA varied from 0.05 to 0.84% of total FA (Figure 1). Based on previous studies the FAs 16:1ω8c, 16:1ω8t, 16:1ω6c, 16:1ω5t, 18:1ω8c and 18:1ω8t are signature FAs for MOB. The total contribution of these FAs was higher in organisms with low δ13C (Table 1, Figure 1). Two discrete groups arose in Figure 1: (A) organisms with a higher total content of MOB signature FAs and lower δ13C values, and (B) organisms with lower content of MOB signature FAs and higher δ13C values. The former group includes Ceratopogonidae and the fish Anadoras grypus with the highest contribution of MOB fatty acids (0.84 and 0.79%, respectively), Chironomidae sp.1 (0.73%), Cyphocharax sp. (0.71%) and the ephemeropteran Campsurus sp. (0.66%). These five organisms were also depleted in 13C (δ13C of -38.1, -36.5, -37.1, -36.1 and -39.7, respectively). All these organisms can feed at oxic-anoxic interfaces where MOB abundance is likely to be high, which provides an explanation to the comparatively high consumption and incorporation of MOB carbon into their biomass. Zooplankton and Chaoborus sp. showed the lowestδ13C values (-42.0 and 40.2, respectively) and a MOB-specific FA contribution of 0.45 and 0.50%, respectively. Calheiros (unpublished data) also found low values ofδ13C in zooplankton (-42.7 to -31.6), benthic chironomids (-62 to -49) and ephemeropterans (-41.4 to -34.3) in another Pantanal lake, suggesting an assimilation of biogenic CH4. Other taxa in group A include the fishes Potamorhina squamoralevis, Crenicichla sp., Leporinus friderici and Steindachnerina brevipinna which had equally lowδ13C values (?13C-36) and a range of MOB-specific FA content from 0.43 to 0.66%. These fishes can all have benthivorous and omnivorous or detritivorous feeding habits, except Crenicichla sp., which is mainly carnivorous.
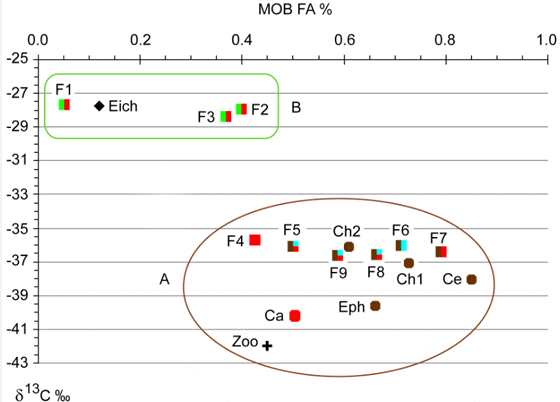
A higher variability in FA composition than in δ13C could be explained by physiological differences among taxa regarding to what extent various FAs are metabolized or stored in muscle tissue. Branched FA are often difficult to metabolize and can therefore be magnified in the tissues. Lipid composition in fish muscle varies mainly with fish diet. The metabolism of total lipids and fatty acids might be related to age, sex, reproductive cycle and capture period of the fish and influenced by environmental factors as seasonal hydrological cycle and food availability. Besides, lipids are unevenly stored among tissue types throughout the year. Differences in migratory behavior and life history may explain distinctive lipid dynamics among fish species. In addition, some organisms might have a more homogeneous diet which could lead to a lower isotopic variation. Calheiros (unpublished data) reported low δ13C values for P. squamoralevis (mean -36.9) and small variations in isotopic signals among seasons, which could be due to a more specialized diet. Wantzen pointed out that seasonal variations in isotopic signal of fish species in Pantanal were more prominent in less specialized omnivores, invertivores and some carnivores, while more specialized detritivores and herbivores appeared to be more influenced by δ13C changes in the diet affected by biogeochemical processes.
Group B (Figure 1), formed by the less 13C-depleted fish species Tetragonopterus argenteus (-27.7), Astyanax cf. bimaculatus (-28.0) and Parauchenipterus galeatus (-28.4), had lower contents of MOB-specific FA (0.05, 0.40 and 0.37%, respectively). The higher δ13C values in combination with lower MOB FA content of these three fishes indicate a lower incorporation of MOB biomass. This conclusion is in accordance with their feeding habits. These taxa are known to be pelagic or living close to aquatic plants, presenting omnivorous habits with tendency to herbivory-invertivory. Wantzen et al. found similar isotopic signatures for T. argenteus and A. bimaculatus in Pantanal. Although aquatic C4 plants were not the major carbon source, they suggested that at least part of the A. bimaculatus diet would be derived from aquatic C4 grasses, which have higher δ13C values than C3 plants and occur in small patches among large mats of C3 macrophytes. The water hyacinth Eichhornia crassipes, a C3 plant that totally dominates the macrophyte community in the studied environment, had a similar δ13C signature (-27.8). Therefore, a combination of carbon sources from C3 plants, C4 plants and MOB could yield a 13C signature similar to C3 plants.
Traces of MOB FA found in plant samples might have two different causes. First, MOB FA in plant samples could be due to the difficulty to completely remove biofilms from the plant surface, especially those attached to the submerged roots. Secondly, there are studies that revealed the presence of MOB inside living tissues of submerged aquatic plants (Sphagnum mosses). Methane-derived carbon was incorporated into plant lipids when mosses were submerged. Even though E. crassipes is a floating macrophyte and not a moss, we cannot exclude the possibility of this phenomenon.
The dendrogram in Figure 2 illustrates the clustering of organisms based on contributions of the different MOB FAs only (i.e. with no consideration to 13C signatures). In contrast to the grouping based on total MOB FA and 13C values illustrated in Figure 1, this clustering approach depends on the relative contribution of the individual MOB FAs, but yields a similar overall result. Four arbitrary cutoff lines, at Bray-Curtis similarities of 0.4, 0.6, 0.8 and 0.87 were used as reference points for identifying clusters.
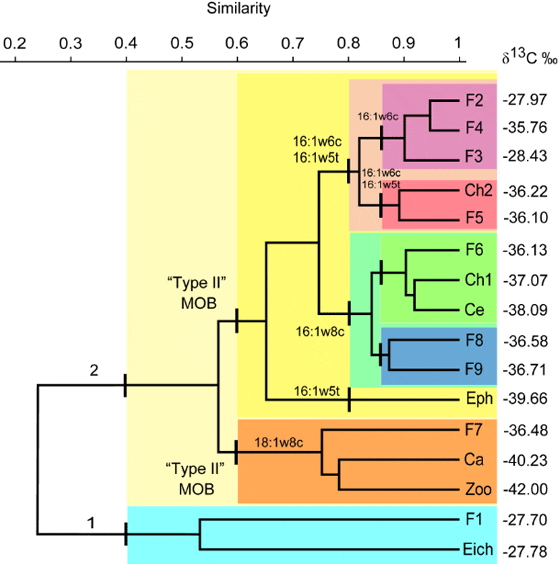
At a similarity of 0.4, two distinct clusters can be seen. Cluster 1 grouped organisms with the lowest contributions of MOB FA, i.e. E. crassipes and T. argenteus. This is in accordance with the fact that T. argenteus is an omnivorous fish with tendency to herbivory-insectivory, feeding mainly on vascular plants, plant debris and terrestrial insects. Cluster 2 was formed by all other organisms with higher MOB FA values.
At a similarity of 0.6, cluster 2 was divided into two new clusters: One had a high contribution of the FA 18:1?8c, which is highly specific for type II MOB, and was therefore named Type II MOB. The other cluster was based on high contribution of specific FA markers for MOB type I, and named Type I MOB. Inside cluster Type II MOB, Chaoborus sp. and zooplankton were grouped together since they presented the highest contributions of 18:1ω8c (84.6% and 68.9%, of the total MOB FA, respectively), followed by A. grypus (56.7% of the total MOB FA). Chaoborid larvae are zooplankton predators, whose preferred food source are microcrustaceans, although they eat a wide variety of animals including dipteran larvae, oligochaetes, rotifers, other chaoborids and even dinoflagellates. It should be noted that zooplankton in this shallow tropical lake are not restricted to autotrophic or heterotrophic production in the water column, but can also access food items from surface sediments which are frequently resuspended, and from the biofilms in the root zone of floating E. crassipes. Anadoras grypus is an invertivore, bottom-feeding fish.
Within cluster Type I MOB, three clusters were identified at a cutoff similarity of 0.8:
- The ephemeropteran Campsurus sp. which showed the highest content of the FA 16:1ω5t. Nymphs of Campsurus are bottom collectors/gatherers and construct their tunnels in soft substrates like mud or sand.
- A cluster of organisms with the highest contributions of 16:1ω8c which are mainly benthivorous and iliophagous (mud eaters; Cyphocharax sp., Chironomidae sp.1, Ceratopogonidae, L. friderici and S. brevipinna). Inside this cluster, two sub-groups emerged: one formed by Cyphocharax sp., Ceratopogonidae and Chironomidae sp.1, and another with L. friderici and S. brevipinna. Species of Cyphocharax pick mainly biofilms from the bottom substrates. Larvae of the aquatic insects Ceratopogonidae and Chironomidae rely on different food sources. Some are detritivorous while others may feed also on periphyton. Both families have collector and scraper feeding habits, live close to the bottom and/or associated to roots of aquatic plants. The last two species L. friderici and S. brevipinna were the most 13C depleted fishes and they have similar feeding habits. Both fishes are iliophagous and feed on detritus, benthic filamentous algae and invertebrates associated to bottom substrates.
- Two groups with high contributions of 16:1ω6c and 16:1ω5t (A. cf. bimaculatus, Crenicichla sp., P. galeatus, Chironomidae sp.2 and P. squamoralevis). The group of Chironomidae sp.2 and P. squamoralevis showed similar proportions of both fatty acids 16:1ω6c and 16:1ω5t, while A. cf. bimaculatus, Crenicichla sp. and P. galeatus were similar to each other in terms of primarily 16:1ω6c. Potamorhina squamoralevis feeds mainly on detritus and benthic algae and invertebrates associated with bottom sediments. Astyanax bimaculatus is an omnivorous fish which consumes mainly insects. Species of Crenicichla are stalking predators, feeding on insects and fishes. The diet of P. galeatus consists mainly of terrestrial and aquatic insects, but also aquatic invertebrates, fishes, fungi, algae, higher plants and detritus.
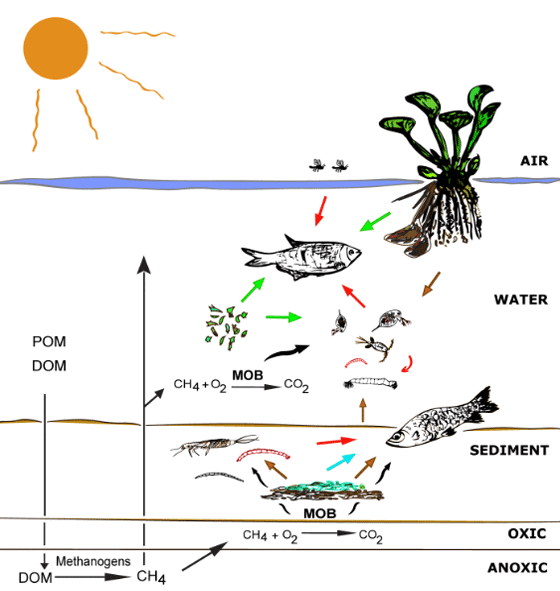
The presence of MOB signature FAs in benthic aquatic insects, planktonic organisms as chaoborids and zooplankton, and fishes showed the importance of biogenic methane to different aquatic compartments in this Pantanal ecosystem (Figure 3). We used a simple two-source mixing model based on 13C signatures to estimate the relative importance of methane-derived carbon as described by Trimmer et al. Assuming E. crassipes and MOB as end members, it was estimated that methane oxidation contributes on average 13% and 1% to the carbon content in fishes from group A and B respectively (Figure 1). However, this value is only an estimate, as the 13C signature of many important carbon sources (phytoplankton, terrestrial DOM from C3 and C4 plants) were not determined and could not be included in this model. The flood pulse also changes the lake dynamics, and may affect the relative importance of MOB for the food chain. We applied the same model for aquatic insects and zooplankton from this lake and obtained a contribution of 16% and 22% as carbon source respectively, a similar value as the results reported by Ravinet et al. and Trimmer et al.
In the present study, the relationship between contribution of MOB FAs and δ13C seems to be driven more by the MOB type II FA (18C) than by MOB Type I FAs (16C). It is possible that MOB type II dominates over type I in this wetland and the Pantanal floodplain in general. Sundh et al. pointed out that most studies in temperate and boreal lakes show a predominance of type I over type II, while a few studies reported dominance of MOB type II in tropical freshwaters (e.g.). Along the same lines, a recent study indicated that type II MOB might drive CH4 oxidation in flood-pulsed wetlands.
The two groups of MOB have different growth characteristics and respond differently to variation in temperature, pH and concentrations of oxygen and methane. Low temperatures seem to favor the development MOB type I over type II. With respect to oxygen and methane supply, MOB type II often dominates at relatively high methane and low oxygen concentrations, while MOB type I dominates at low CH4 and high oxygen concentrations.
The Pantanal floodplain is characterized by conditions that together favor production and emissions of CH4, such as high primary productivity, shallow depth of inundation, and high temperature. Such conditions could be expected to be favorable for aerobic methane oxidizers with a probable selection for type II, in line with our results. However, although FAs of MOB type II showed a stronger correlation with 13C signatures, we have to be careful to draw the conclusion that MOB type II dominates fish consumption of MOB in Pantanal as the actual levels of MOB type I and type II FAs in the fishes were overall quite similar.
This study showed, by combining two independent tracers (MOB FA and δ13C), that MOB carbon was transferred through the food web up to the fish level. Hence, our data demonstrate that CH4 can be a significant carbon source not only for the microbial food web and invertebrates, but also for higher trophic levels. The Pantanal region is characterized by high CH4 production and emissions, and transfer of CH4 carbon throughout the food web to various fish species could be a common phenomenon in CH4-rich ecosystems.
December 2012
