The aim of this study was to analyse the variation of biochemical and molecular characteristics of Y. ruckeri isolates collected in north west Germany as a basis for strain differentiation. The isolates originated mainly from rainbow trout and were characterised by biochemical profiling, 16S rDNA sequencing, repetitive sequence-based PCRs, including (GTG)5-PCR, BOX-PCR, ERIC-PCR and REP-PCR, and pulsed-field gel electrophoresis (PFGE).
Results
Biochemical and physiological characterisation
Among the variable biochemical characteristics of all 83 Y. ruckeri isolates (Table 1), gelatine hydrolysis, Voges- Proskauer (VP) reaction and Tween hydrolysis were the most variable biochemical reactions. The results of VP, nitrate production, citrate utilization and gelatine hydrolysis from both, conventional biochemical analysis and testing by the API 20E system, were listed also in Table 1. For motility, Tween 80/20 hydrolysis and citrate utilization, the isolates obtained in Lower Saxony (LS) from fish species other than rainbow trout were more similar to the reference strain DSM 18,506 than the isolates obtained from rainbow trout hatcheries in NRW and LS. Only 16 isolates hydrolysed Tween 80/ 20. For these isolates motility was confirmed by microscopical inspection as well as by cultivation in API M medium. Those isolates were recognized as non-motile isolates. In particular, isolates collected from trout hatcheries in NRW were lacking flagella and motility; 92 per cent of the isolates from this federal state were non-motile (Table 1).
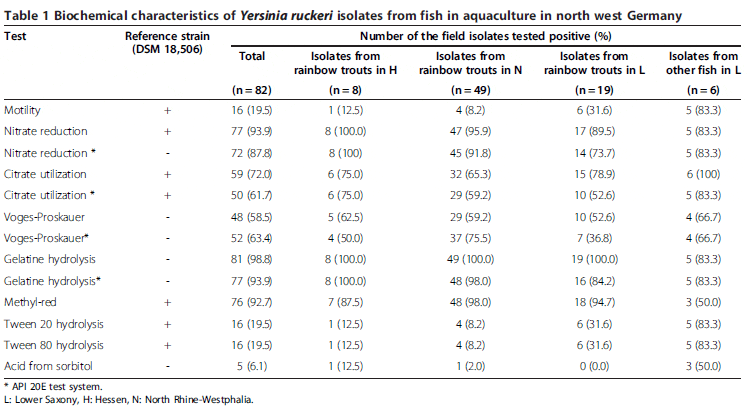
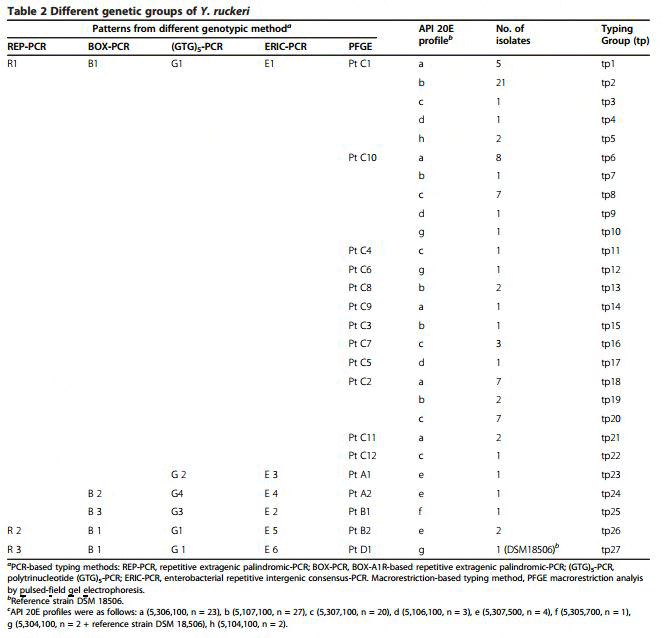
In the API 20E test, 27 isolates showed the numeric profile 5,107,100, 23 isolates with the numeric profile 5,306,100. These profiles differed from that of the reference strain DSM 18,506 with the profile 5,304,100. For a biochemical typing of the isolates, typing numbers from “a” to “g” were assigned to individual isolates according to their profile number (Table 2).
Genetic characterisation
When the isolates were analysed by repetitive sequencebased PCR assays, two amplicon patterns were obtained after REP-PCR, five patterns by ERIC-PCR, four patterns by (GTG)5-PCR and three patterns by BOX-PCR (Figure 1). The similarity rates in REP-PCR were clearly lower than in the other repetitive sequence-based PCRs. In BOX-PCR and (GTG)5-PCR, the reference strain DSM 18,506 exhibited the same amplicon patterns as most of the isolates (Figure 1). By REP-PCR, 3 or 19 bands were seen in the size range of 200–2000 bp; by BOX-PCR, 18–19 bands were found distributed between 340–1550 bp; by (GTG)5-PCR, 11–13 bands were detected from 390–2500 bp; and by ERIC-PCR, 20–25 bands were present in the range from 200–2000 bp (Figure 1). Five isolates, which were positive in sorbitol fermentation, did not exhibit the 430 bp amplicon in ERIC-PCR. Only five isolates from fish species other than rainbow trout showed some differences in repetitive PCRs. These five isolates were additionally confirmed as Y. ruckeri (identities = 100 per cent, data not shown) by 16S rDNA sequencing; while the nucleotide sequence of the reference strain DSM 18,506 was not completely identical to the sequence reported from the strain ATCC 29,473 (identities = 99 per cent, data not shown). Conversely, all the isolates from rainbow trout showed rather uniform patterns of repetitive sequence-based PCR.

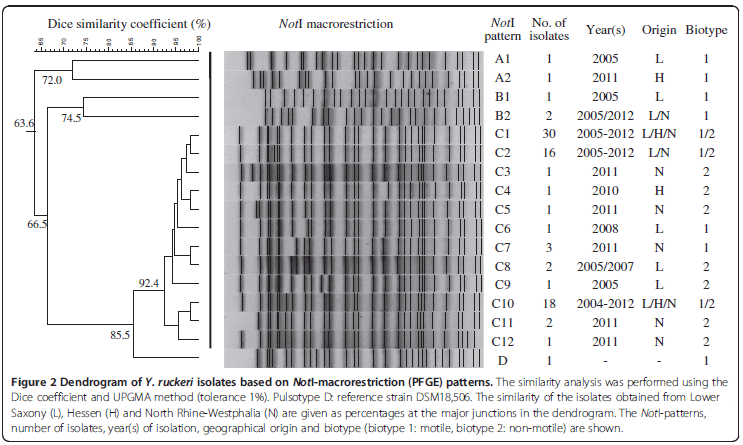
NotI-directed PFGE yielded four major PFGE groups A-D. A total of 17 different PFGE types (A1, A2, B1, B2, C1-C12, D) were detected among the 83 Y. ruckeri isolates examined (Figure 2, Table 2). Cluster analysis of these PFGE types showed that the isolates from north west Germany were highly uniform, whereby 30 isolates (36.1 per cent) belonged to PFGE type C1 and 18 isolates to PFGE type C10 (Figure 2). Compared with the reference strain DSM 18,506, all the isolates were different. The isolates from rainbow trout were clustered together (PFGE group C). Isolates from pike, koi and brown trout (PFGE groups A and B) showed the greatest variation when compared to rainbow trout isolates. All isolates previously assigned to biotype 2 clustered in PFGE group C. Isolates from this PFGE group were found in the federal states of LS, Hessen (H) and NRW (Figure 2). Most of the isolates were distributed in all three federal states (PFGE type C1 and C10), while isolates of PFGE type C2 were found only in LS and NRW (Figure 2).
Discriminatory indices of the typing methods applied
As a discriminatory index, Simpson’s index of diversity was calculated for each typing method used to differentiate all Y. ruckeri isolates. Discriminatory indices varied distinctly between the different typing methods. REPPCR had the lowest D value of 0.048. This means that two randomly selected isolates of this test population had a probability of 4.8 per cent of showing a different REP-PCR pattern. Slightly higher D values were calculated for BOXPCR (D = 0.071), (GTG)5-PCR (D = 0.095) and ERIC-PCR (D = 0.140). Highest D values were seen for biotyping via API 20E (D = 0.763) and NotI-directed PFGE (D = 0790). According to their biochemical and molecular characteristics as obtained by testing with API 20E, PFGE and various PCR-based methods, the Y. ruckeri isolates from north west Germany could be subdivided into 27 different groups (Table 2). Most of the isolates from north west Germany (30, 36.6 per cent) belonged to the group (R1/B1/G1/E1/PFGE type C1). Additional information about the origins and isolated years were shown in Additional file 1: Table S1.
Discussion
Since the first isolation of Y. ruckeri in Idaho, USA in 1950s, outbreaks of ERM were frequently reported, especially from the aquaculture industry in countries such as Croatia, France, Finland, Germany, Italy, Switzerland and the UK. The bacterium was isolated from different host species, including several salmonid species, channel catfish, goldfish, common carp, European eel, coalfish, and perch. Typing of Y. ruckeri isolates from various geographic locations or different fish species has been performed for taxonomic or epidemiological purposes and initially it was based on phenotypical characteristics including the results of biochemical and serological tests. In addition, molecular techniques, including random amplification of polymorphic DNA (RAPD), multilocus sequence typing (MLST) or pulsed-field gel electrophoresis (PFGE) have been applied to study the genetic diversity of Y. ruckeri isolates. In a study on isolates from various geographic origins, including Chile, Peru, USA, UK, and mainland Europe, a considerable intraspecies diversity of Y. ruckeri was found on the basis of the phenotypical characteristics obtained from API 20E, lipopolysaccharide (LPS) and outer membrane profiles as well as by genetic analysis using MLST profiles.
The isolates from north west Germany investigated in the present study, showed a strong homogeneity of biochemical results and API 20E profiles with some variability in gelatine hydrolysis, VP reaction and Tween hydrolysis. Variation in some biochemical tests for Y. ruckeri has been reported in previous studies, but this variation could not be used as a distinguishing characteristic for phenotypical traits such as serotype or biotype. While the majority of Y. ruckeri isolates from farmed rainbow trout in Peru or Atlantic salmon in Chile were motile, only 16 out of 82 isolates from the present study belonged to biotype 1 (motile and lipase positive). All other isolates could not hydrolyse Tween 80/20, were lipase-negative and lacked flagella.
The API 20E profiles 5,306,100, 5,107,100, 5,104,100 were previously obtained from Y. ruckeri isolates from southwest Germany, the profiles 5,307,100 and 5,304,100 were already reported from Spanish isolates. While the profiles 5,106,100 and 5,307,500 were also previously reported from Y. ruckeri isolates, there was no record about an isolate with the numeric profile 5,305,700. However, 16S rRNA sequencing confirmed this isolate as Y. ruckeri (identity = 100 per cent, data not shown).
In addition to the biochemical characteristics, some genetic variability was observed in isolates using several repetitive sequence-based PCR techniques including BOX-PCR, ERIC-PCR, (GTG)5-PCR, REP-PCR as well as pulsed-field gel electrophoresis. An analysis of the isolates from north west Germany by repetitive sequence based PCRs gave 3 to 6 groups, with the majority of isolates in one group. Similar observations were already reported from Latin American isolates. ERIC-PCR showed the highest discriminatory power. While in previous studies both ERIC-PCR and REP-PCR were considered of some value as tools to study ERM epidemiology, in our study, REP-PCR showed the lowest discriminatory power and could not differentiate the isolates very clearly. This could depend on the different origin of the isolates: our isolates originated mainly from rainbow trout, while Bastardo et al. studied isolates from Atlantic salmon. When Y. ruckeri isolates from Peruvian rainbow trout were analysed by REP-PCR, more homogeneous amplicon patterns were observed as well.
PFGE has been frequently applied for detailed molecular typing of different bacteria, as well as for a differentiation among isolates of Y. ruckeri. In the present study, the typing results by PFGE showed strong homogeneity among the isolates obtained from farmed fish species in north west Germany. More than half of the Y. ruckeri isolates belonged to the two PFGE types C1 and C10, with 36.1 per cent of the isolates sharing the same PFGE type C1. These two PFGE types belong to the group C PFGE cluster, which comprises isolates from rainbow trout and represents the majority of the isolates. The isolates showing PFGE types A1/A2 or B1/B2 also differed from the other isolates in at least some of their repetitive sequences-based PCR patterns (Table 2). In the present study, no distinct differences were observed in PFGE types from biotype 1 and biotype 2 isolates from rainbow trout.
The data presented in this study may suggest that the isolates from north west Germany originated from the same ancestor and could indicate a clonal population structure. In a previous study on the genetic diversity of Y. ruckeri isolates, analysed by MLST, a large number of isolates also belonged to just two sequence types (ST), which formed one clonal complex with 21 out of 30 recognised STs. However, the MLST analysis revealed that genetic recombination appeared to play a greater role than mutation for the generation and maintenance of genetic diversity within the population of Y. ruckeri. This would be contradictory to the concept of clonal expansion of a population from a common ancestor that diversified mainly by mutation rather than recombination. MLST results from Y. ruckeri rather suggest an epidemic population structure, as it was observed for instance for Vibrio parahaemolyticus. The epidemiology of Yersinia ruckeri, which started as a geographically isolated disease and quickly became disseminated, also was considered to support the epidemic model.
Pulsed-field gel electrophoretic analysis of Y. ruckeri isolates resulted in a discriminatory index (D) of 0.790, whereas repetitive sequence-based PCR approaches yielded low D values between 0.048 and 0.140. As the D value describes the probability that two randomly selected isolates show different profiles by the method applied, methods with low D values are considered not to be sufficiently discriminative for the detection of differences among Y. ruckeri isolates. In a previous analysis, “medium to high” D values ranging between 0.71–0.74 were obtained by the API 20E, lipopolysaccharide and outer membrane protein analysis, while ERIC-and REP-PCR were valuable in combination with other techniques. Similar results were reported from Y. ruckeri isolated in Turkey. The combination of several typing methods, as performed by Bastardo et al. showed that isolates with similar characteristics were associated with certain fish species and/or predominated in some geographical areas. In the present study, the combination of six typing methods, including API 20E, PFGE and four different repetitive sequence PCR techniques facilitated the identification of 27 different groups. With this set of methods, most of the isolates from rainbow trout (97.3 per cent) could be differentiated from the isolates obtained from pike, koi and brown trout. Biotype 2 isolates were only detected in rainbow trout in this study. The genomic variation of Y. ruckeri isolates provides information for comparing their distribution and the relationships among individual isolates from different regions in north west Germany.
Conclusions
Most of the isolates from north west Germany belonged to biotype 2. The combination of different typing methods could well differentiate Y. ruckeri isolates. The results of this study showed the diversity of isolates present in north west Germany. Moreover, these data enable the identification and in-depth characterisation of pathogenic Y. ruckeri isolates. As a consequence of this detailed characterisation, these data can be used to examine the distribution of these isolates in the field and may be used as a basis for preventive disease monitoring plans.
November 2013

